Nová studie spojuje drobné odchylky v pohybu s rizikem duševních potíží
30. 07. 2026
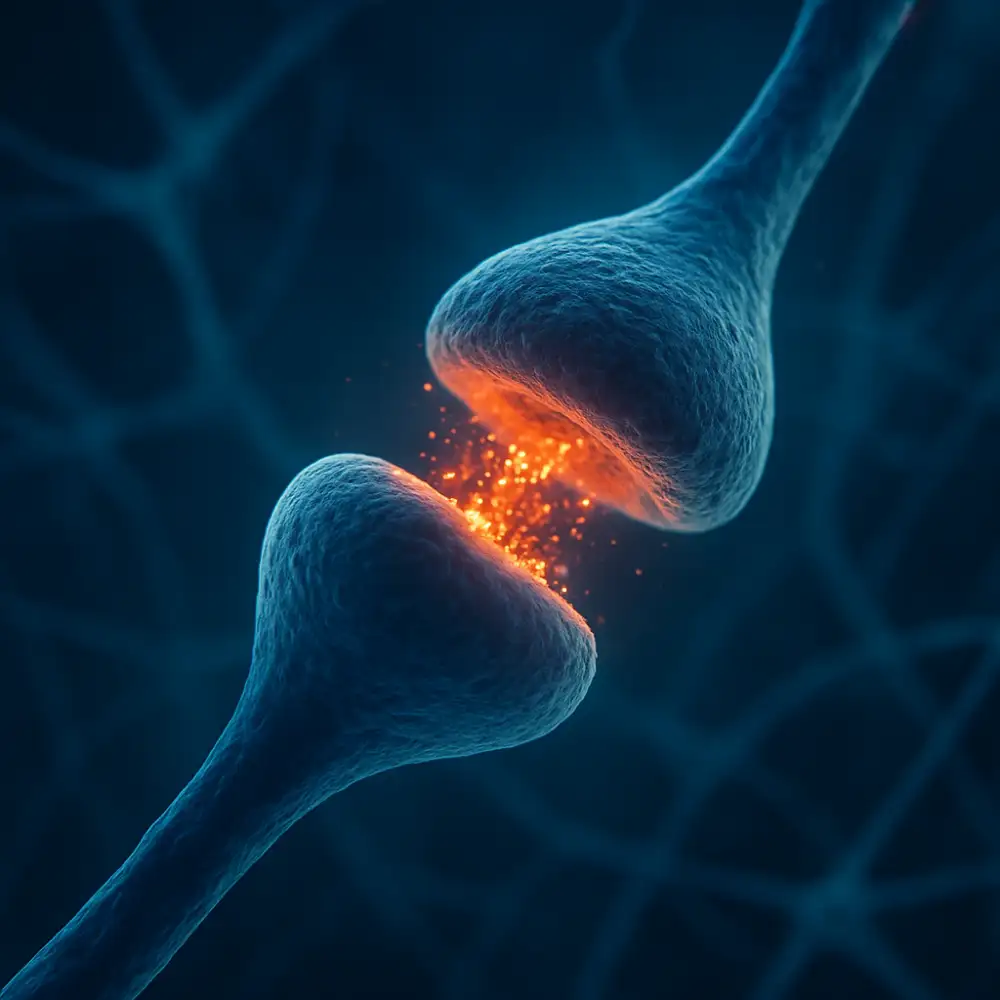
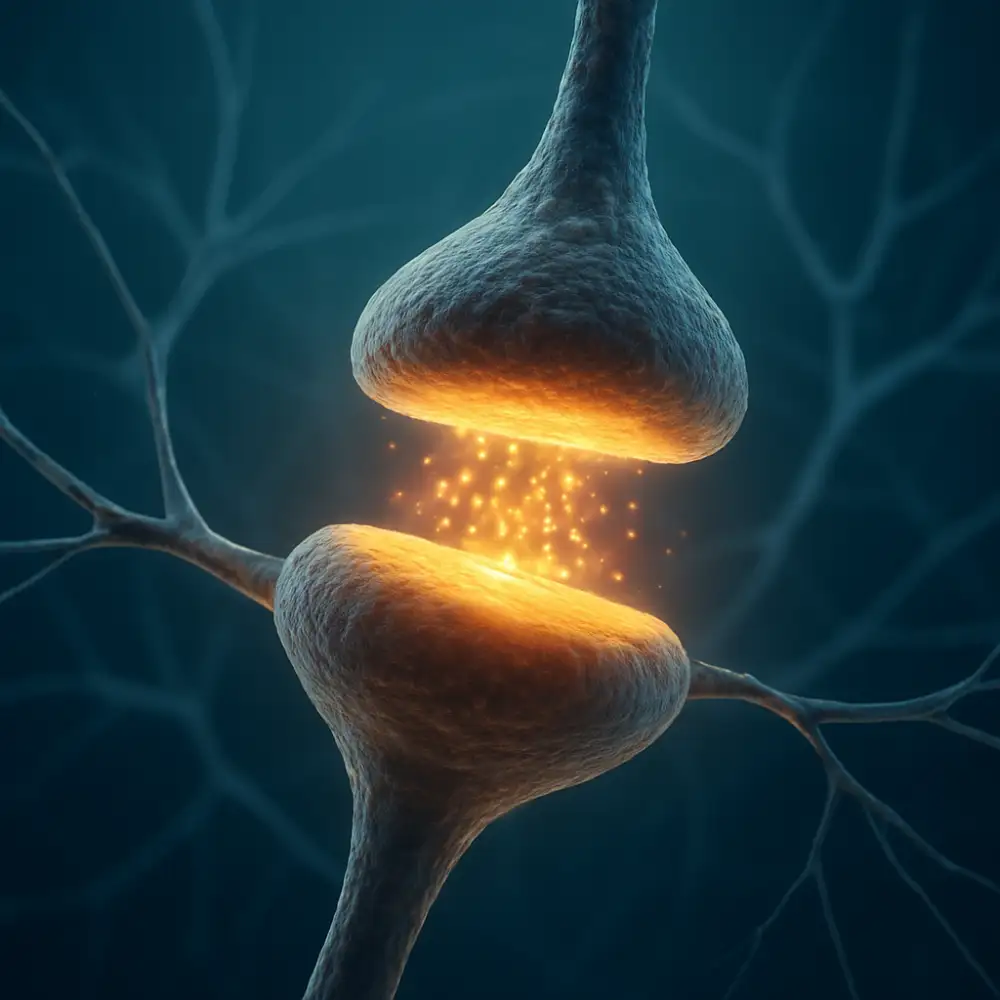
Synapse představuje jednu z nejzásadnějších struktur celého nervového systému, bez níž by přenos informací mezi neurony nebyl vůbec možný. Jedná se o specializované místo kontaktu mezi dvěma nervovými buňkami, případně mezi neuronem a efektorovou buňkou, jako je svalová nebo žlázová buňka. Synapse umožňuje přenos nervového vzruchu z jednoho neuronu na druhý, přičemž tento proces probíhá buď prostřednictvím chemických látek zvaných neurotransmitery, nebo přímým elektrickým spojením. Pochopení toho, co synapse je a jak funguje, je naprosto klíčové pro porozumění celé řadě neurologických i psychiatrických onemocnění, ale také pro základní chápání toho, jak mozek zpracovává informace, jak se učíme a jak si pamatujeme.
Z anatomického hlediska se synapse skládá ze tří základních částí. Presynaptická část odpovídá zakončení axonu vysílajícího neuronu, kde jsou uloženy synaptické váčky naplněné neurotransmitery. Synaptická štěrbina pak představuje úzký prostor mezi oběma buňkami, do nějž jsou tyto látky uvolňovány. Postsynaptická membrána přijímajícího neuronu nese receptory, které jsou schopny rozpoznat konkrétní neurotransmitery a reagovat na jejich přítomnost. Celý tento systém funguje s obdivuhodnou přesností a rychlostí, přičemž přenos signálu přes synaptickou štěrbinu trvá pouhé milisekundy.
Role synapse v nervovém systému je nesmírně komplexní a mnohostranná. Není to jen prosté předávání elektrického signálu, jak by se mohlo na první pohled zdát. Synapse je místem, kde dochází k modulaci, zesílení nebo naopak utlumení nervového signálu, čímž se celý nervový systém stává neobyčejně flexibilním a adaptabilním nástrojem. Jeden neuron může být spojen tisíci synapsemi s dalšími neurony, přičemž každé takové spojení může mít excitační nebo inhibiční charakter. Výsledný efekt na postsynaptický neuron pak závisí na součtu všech těchto vlivů, což je princip nazývaný synaptická integrace.
Zvláštní pozornost si zaslouží schopnost synapsí měnit svou sílu v závislosti na předchozí aktivitě, tedy fenomén označovaný jako synaptická plasticita. Právě tato vlastnost stojí za naší schopností učit se novým věcem a uchovávat vzpomínky. Dlouhodobé posílení synaptického spojení, anglicky označované jako long-term potentiation, je považováno za jeden z hlavních buněčných mechanismů paměti. Naopak dlouhodobé oslabení synapse hraje roli v zapomínání a přeorganizování nervových okruhů.
Neurony komunikují výhradně prostřednictvím synapsí, a proto jakékoli narušení synaptického přenosu má přímý dopad na funkci celého nervového systému. Poruchy v synaptické transmisi jsou spojovány s celou řadou závažných onemocnění, včetně Alzheimerovy choroby, schizofrenie, deprese, epilepsie nebo Parkinsonovy nemoci. Mnoho léků, které se v současnosti používají k léčbě těchto stavů, působí právě na úrovni synapse, ať už ovlivňují uvolňování neurotransmiterů, blokují jejich receptory nebo brání jejich zpětnému vstřebávání.
Synapse tedy není pouhým pasivním přechodem mezi dvěma neurony, ale dynamickým, vysoce regulovaným rozhraním, které určuje kvalitu a charakter veškeré nervové komunikace. Její správná funkce je podmínkou nejen pro fyzické pohyby a smyslové vnímání, ale také pro emoce, myšlení, rozhodování a veškeré kognitivní procesy, které nás definují jako myslící bytosti. Pochopení synapse na molekulární úrovni otevírá dveře k novým terapeutickým přístupům a hlubšímu poznání toho, jak lidský mozek skutečně funguje.
Každý neuron v nervové soustavě je jedinečná buňka, která disponuje specifickou architekturou přizpůsobenou přenosu a zpracování informací. Abychom pochopili, jak synapse funguje, je nezbytné nejprve porozumět tomu, jak jsou uspořádány neurony, které ji tvoří. Synapse není izolovaný bod kontaktu – je to výsledek precizní spolupráce dvou odlišně specializovaných zakončení, přičemž každé z nich plní zcela jinou úlohu.
Presynaptický neuron je ten, který signál vysílá. Jeho tělo, označované jako soma, obsahuje jádro s genetickou informací a veškeré organely nezbytné pro metabolismus buňky. Z těla neuronu vybíhají dendrity, které přijímají podněty z okolních neuronů, a jeden axon, jenž slouží k vedení elektrického impulzu směrem od těla buňky k cílovému místu. Axon může být ovinut myelinovou pochvou, která dramaticky zvyšuje rychlost vedení vzruchu díky takzvanému saltatornímu vedení, při němž elektrický signál přeskakuje mezi Ranvierovými zářezy. Na konci axonu se nachází axonální terminála, neboli presynaptické zakončení, které je přímo zodpovědné za uvolňování neurotransmiterů do synaptické štěrbiny.
Uvnitř presynaptického zakončení se nacházejí synaptické váčky – malé membránové struktury naplněné neurotransmitery. Tyto váčky jsou připraveny k fúzi s presynaptickou membránou v okamžiku, kdy do zakončení dorazí akční potenciál. Spolu s váčky jsou zde přítomny mitochondrie, které zajišťují energetický zásobník ve formě ATP, nezbytný pro syntézu a recyklaci neurotransmiterů. Presynaptická membrána obsahuje napěťově řízené vápníkové kanály, jejichž otevření po příchodu akčního potenciálu umožní příliv vápenatých iontů, a právě tento příliv spouští exocytózu synaptických váčků.
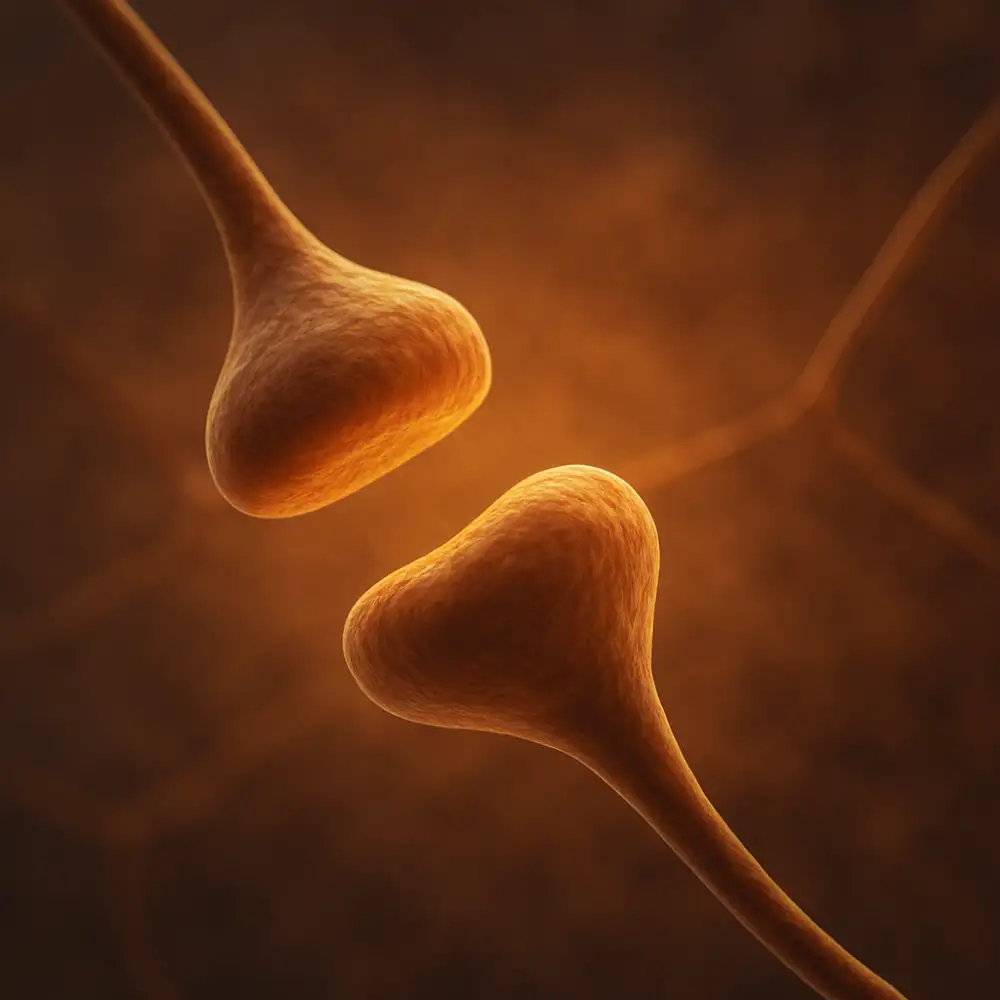
Postsynaptický neuron naproti tomu signál přijímá. Jeho membrána v oblasti synaptického kontaktu, označovaná jako postsynaptická densita, je obohacena o velké množství receptorů specificky vázajících uvolněné neurotransmitery. Tyto receptory mohou být ionotropní, tedy přímo spojené s iontovými kanály, nebo metabotropní, které svůj účinek zprostředkovávají prostřednictvím druhých poslů a G-proteinů. Hustota a typ těchto receptorů určuje, jak citlivě a jakým způsobem postsynaptický neuron na přijatý signál zareaguje.
Mezi presynaptickým zakončením a postsynaptickou membránou se nachází synaptická štěrbina, úzký prostor o šířce přibližně 20 až 40 nanometrů. Tento prostor není prázdný – je vyplněn extracelulární matrix a různými proteiny, které pomáhají udržovat strukturální integritu synapse a modulují difuzi neurotransmiterů. Rychlost, s jakou neurotransmiter překoná tuto vzdálenost, je překvapivě vysoká, přesto je synaptický přenos pomalejší než čistě elektrické vedení, což má svůj fyziologický smysl.
Celá tato architektura není statická. Synapse se neustále přestavují, posilují nebo zeslabují v závislosti na aktivitě neuronu, a právě tato plasticita je základem učení a paměti. Presynaptický neuron může měnit množství uvolňovaných neurotransmiterů, postsynaptický neuron zase počet a citlivost svých receptorů. Tento dynamický vztah mezi oběma neurony tvoří základ toho, čemu říkáme synaptická plasticita, a bez ní by nebylo možné žádné přizpůsobování nervové soustavy novým podmínkám ani ukládání zkušeností do dlouhodobé paměti.
Synapse představují klíčové struktury nervového systému, jejichž prostřednictvím dochází k předávání informací mezi neurony. Způsob, jakým je signál přenášen z jednoho neuronu na druhý, určuje základní klasifikaci synapsí do několika kategorií, přičemž každá z nich má své specifické vlastnosti, funkce a místo v celkové architektuře nervových okruhů.
Nejrozšířenějším typem jsou chemické synapse, které tvoří naprostou většinu synaptických spojení v lidském nervovém systému. Jejich princip spočívá v tom, že elektrický signál přicházející ve formě akčního potenciálu do presynaptického zakončení vyvolá uvolnění chemických látek nazývaných neurotransmitery. Tyto látky jsou skladovány ve specializovaných váčcích – synaptických vezikulách – a jejich uvolnění do synaptické štěrbiny je závislé na přítoku vápenatých iontů, který nastane v okamžiku depolarizace membrány. Neurotransmitery následně difundují přes synaptickou štěrbinu a váží se na specifické receptory postsynaptické membrány, čímž vyvolávají změny v elektrickém potenciálu postsynaptického neuronu nebo spouštějí různé intracelulární signální kaskády. Celý tento proces trvá řádově milisekundy, přičemž zpoždění způsobené chemickým přenosem je jednou z charakteristických vlastností tohoto typu synapsí.
Chemické synapse lze dále rozdělit podle účinku, který mají na postsynaptický neuron. Excitační synapse zvyšují pravděpodobnost vzniku akčního potenciálu v postsynaptickém neuronu tím, že způsobují jeho depolarizaci. Typickým excitačním neurotransmiterem je glutamát, který se váže na různé typy receptorů, například AMPA nebo NMDA receptory. Naproti tomu inhibiční synapse snižují excitabilitu postsynaptického neuronu a brání vzniku akčního potenciálu. Hlavním inhibičním neurotransmiterem v centrálním nervovém systému je kyselina gama-aminomáselná, zkráceně GABA, zatímco v periferním nervovém systému tuto roli zastává glycin. Rovnováha mezi excitačními a inhibičními synapsemi je naprosto zásadní pro správné fungování nervových okruhů a jakákoli její porucha může vést k závažným neurologickým nebo psychiatrickým onemocněním.
Zcela odlišný princip přenosu signálu využívají elektrické synapse, které jsou v lidském nervovém systému méně časté, avšak v určitých oblastech a situacích hrají nezastupitelnou roli. Tyto synapse jsou tvořeny specializovanými mezibuněčnými kanály nazývanými gap junctions, neboli nexusy, které přímo propojují cytoplazmu dvou sousedních neuronů. Prostřednictvím těchto kanálů mohou ionty a malé molekuly volně přecházet z jedné buňky do druhé, čímž dochází k přímému šíření elektrického signálu bez nutnosti jakéhokoli chemického zprostředkovatele. Přenos je prakticky okamžitý a na rozdíl od chemických synapsí probíhá obvykle obousměrně, ačkoli existují i elektrické synapse s usměrňovací funkcí, které umožňují tok signálu pouze jedním směrem. Elektrické synapse jsou typické například pro srdeční sval, kde zajišťují synchronizované stahování buněk, ale nacházejí se také v různých oblastech mozku, kde koordinují aktivitu skupin neuronů.
Zvláštní kategorii tvoří smíšené synapse, které kombinují vlastnosti chemického i elektrického přenosu. V těchto synapsích jsou přítomny jak gap junctions, tak i oblasti typické pro chemické synapse s vezikulami obsahujícími neurotransmitery. Smíšené synapse byly popsány například v mozkovém kmeni a v míše a jejich funkce spočívá pravděpodobně v tom, že elektrická složka zajišťuje rychlý přenos základní informace, zatímco chemická složka umožňuje modulaci a přizpůsobení přenosu aktuálním potřebám organismu.
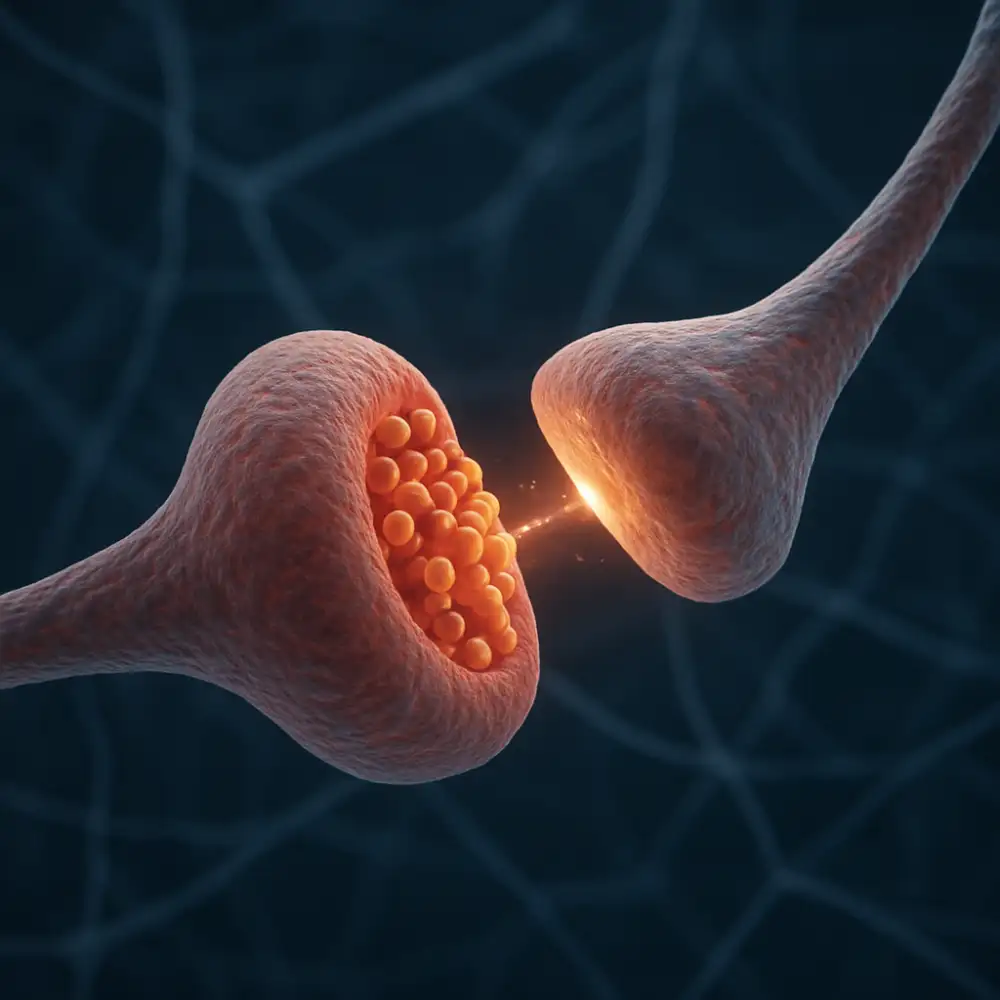
Kromě těchto základních kategorií existují také neuromodulační synapse, jejichž úloha není primárně v rychlém přenosu informace, ale spíše v dlouhodobé modulaci aktivity neuronálních sítí. Tyto synapse uvolňují neuromodulátory, jako jsou dopamin, serotonin, noradrenalin nebo acetylcholin, které mohou působit na vzdálené neurony a ovlivňovat jejich citlivost na jiné synaptické vstupy. Neuromodulace je klíčovým mechanismem, který stojí za procesy jako jsou učení, paměť, pozornost nebo regulace nálady. Poruchy neuromodulačních systémů jsou spojeny s celou řadou onemocnění, včetně Parkinsonovy choroby, deprese nebo schizofrenie.
Pochopení různých typů synapsí a jejich způsobů přenosu signálu je tedy naprosto zásadní nejen pro základní neurovědy, ale také pro klinickou medicínu a farmakologii, neboť naprostá většina léků ovlivňujících nervový systém působí právě na úrovni synaptického přenosu.
Chemické synapsy představují nejrozšířenější typ synaptického spojení v nervové soustavě a jejich fungování je naprosto klíčové pro přenos informací mezi neurony. Na rozdíl od elektrických synapsí, kde dochází k přímému šíření elektrického signálu skrze spojovací kanály, chemické synapsy využívají jako prostředníka speciální molekuly označované jako neurotransmitery. Tento způsob přenosu je sice pomalejší, ale umožňuje mnohem větší flexibilitu a regulovatelnost signálů, což je pro složité funkce mozku naprosto nezbytné.
| Vlastnost | Chemická synapse | Elektrická synapse | Nervosvalová ploténka |
|---|---|---|---|
| Šířka synaptické štěrbiny | 20–40 nm | 2–4 nm | 50–70 nm |
| Rychlost přenosu signálu | 0,5–5 ms | 0,1–0,2 ms | 1–2 ms |
| Typ přenosu | Jednosměrný (chemický) | Obousměrný (elektrický) | Jednosměrný (chemický) |
| Hlavní neurotransmiter | Glutamát, GABA, dopamin | Ionty (Na⁺, K⁺, Ca²⁺) | Acetylcholin |
| Počet v lidském mozku | ~100 bilionů | ~10 bilionů | ~650 milionů |
| Průměr synaptického váčku | 40–50 nm | Není přítomen | 40–60 nm |
| Klidový membránový potenciál neuronu | −70 mV | −70 mV | −90 mV |
| Akční potenciál (amplituda) | +40 mV | +40 mV | +30 mV |
| Plasticita (schopnost změny) | Vysoká (LTP, LTD) | Nízká | Střední |
| Výskyt v CNS | Mozek, mícha | Mozeček, sítnice | Periferní nervový systém |
Celý proces začíná v okamžiku, kdy akční potenciál dorazí do presynaptického zakončení neuronu. Tato oblast, nazývaná také presynaptická terminála nebo synaptický knoflík, je naplněna malými váčky obklopenými membránou, které se označují jako synaptické vezikuly. Každá z těchto vezikulí v sobě ukrývá tisíce molekul neurotransmiteru, připravených k uvolnění ve správný okamžik. Jakmile akční potenciál dosáhne terminály, dojde k depolarizaci membrány a k otevření napěťově řízených vápníkových kanálů. Vápníkové ionty pak proudí z extracelulárního prostoru dovnitř buňky, protože jejich koncentrace je venku podstatně vyšší než uvnitř.
Právě přítok vápenatých iontů spouští kaskádu molekulárních událostí, která vede k fúzi synaptických vezikulí s presynaptickou membránou. Tento děj je zprostředkován skupinou proteinů označovaných jako SNARE proteiny, které fungují jako jakési molekulární spony, jež přitahují vezikulu k membráně a umožňují jejich splývání. Po fúzi se obsah vezikuly vylije do synaptické štěrbiny, tedy do úzkého prostoru mezi presynaptickým a postsynaptickým neuronem. Šířka této štěrbiny se pohybuje přibližně kolem dvaceti až čtyřiceti nanometrů, což je vzdálenost zdánlivě nepatrná, ale pro správné fungování přenosu zcela zásadní.
Neurotransmitery se poté difúzí šíří přes synaptickou štěrbinu a váží se na specifické receptory umístěné na postsynaptické membráně. Tyto receptory jsou vysoce selektivní a reagují pouze na určité typy molekul, což zajišťuje přesnost a specificitu přenosu. Existují dva základní typy postsynaptických receptorů. Ionotropní receptory jsou přímo spojeny s iontovými kanály a jejich aktivace vede k okamžitému otevření kanálu a změně membránového potenciálu postsynaptického neuronu. Metabotropní receptory naproti tomu působí prostřednictvím druhých poslů a jejich efekt je pomalejší, ale déletrvající a může ovlivňovat nejrůznější intracelulární procesy.
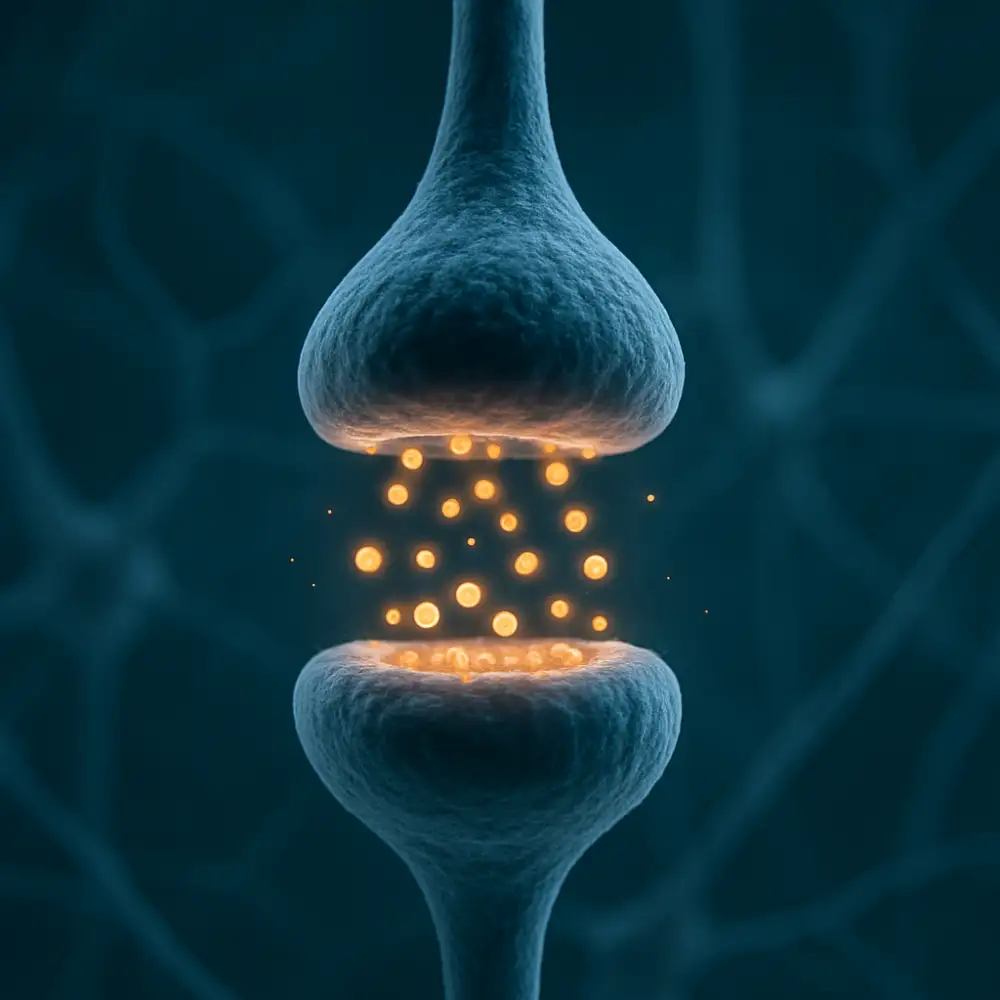
Výsledkem vazby neurotransmiteru na receptor může být buď excitace nebo inhibice postsynaptického neuronu. Excitační neurotransmitery, jako je například glutamát, způsobují depolarizaci postsynaptické membrány a zvyšují pravděpodobnost vzniku nového akčního potenciálu. Inhibiční neurotransmitery, mezi něž patří zejména kyselina gama-aminomáselná, známá jako GABA, naopak způsobují hyperpolarizaci membrány a snižují excitabilitu neuronu. Rovnováha mezi excitačními a inhibičními vlivy je pro správné fungování nervové soustavy naprosto nepostradatelná a její narušení se podílí na vzniku celé řady neurologických a psychiatrických onemocnění.
Po uvolnění a navázání na receptory musí být neurotransmitery z synaptické štěrbiny odstraněny, aby byl přenos signálu přesně časově ohraničen a aby nedocházelo k přetrvávající aktivaci postsynaptických receptorů. K tomu dochází několika mechanismy. Část molekul je zpětně pohlcena do presynaptického zakončení prostřednictvím specifických transportních proteinů, kde mohou být znovu zabaleny do vezikulí a použity při dalším přenosu. Tento proces se nazývá zpětné vychytávání neboli reuptake a je cílem mnoha farmakologicky aktivních látek, včetně antidepresiv ze skupiny selektivních inhibitorů zpětného vychytávání serotoninu. Jiné neurotransmitery jsou enzymaticky štěpeny přímo v synaptické štěrbině, jako je tomu například u acetylcholinu, který je rozkládán enzymem acetylcholinesterázou.
Celý systém chemické synaptické transmise je navíc podroben složité regulaci, která zahrnuje například presynaptické autoreceptory, jež snímají koncentraci neurotransmiteru ve štěrbině a zpětnovazebně regulují jeho další uvolňování. Synaptický přenos tak není jednosměrným procesem, ale dynamickým dialogem mezi oběma neurony, který se neustále přizpůsobuje aktuálním potřebám nervové soustavy. Tato plasticita synaptického přenosu je základem učení a paměti a představuje jeden z nejfascinujících aspektů fungování lidského mozku.
Elektrické synapsy představují fascinující a poněkud opomíjenou kategorii synaptických spojení, která fungují na zcela odlišném principu než jejich chemické protějšky. Zatímco chemické synapsy využívají neurotransmitery jako prostředníky přenosu signálu, elektrické synapsy umožňují přímý tok iontů mezi dvěma neurony, čímž dosahují mimořádně rychlého a spolehlivého přenosu informace. Tento typ spojení byl po dlouhá desetiletí považován za méně významný nebo dokonce primitivní, avšak moderní neurověda odhaluje jeho nezastupitelnou roli v celé řadě fyziologických procesů.
Základem elektrické synapsy jsou specializované struktury zvané gap junctions, neboli spojovací kanálky. Tyto struktury tvoří přímé cytoplazmatické mosty mezi sousedními neurony a jsou sestaveny z proteinů nazývaných konexiny. Každý kanálek se skládá ze dvou hemikanálků, přičemž každý hemikanálek pochází z jednoho z propojených neuronů. Šest konexinových podjednotek se uspořádá do prstencové struktury zvané konexon, a právě dva konexony z protilehlých buněk se setkají a vytvoří funkční kanálek prostupující oběma buněčnými membránami. Průměr tohoto kanálku je dostatečně velký, aby jím mohly procházet nejen ionty, ale i malé molekuly jako cAMP nebo IP3, což dává elektrickým synapsím ještě širší funkční potenciál.
Přenos signálu přes elektrickou synapsi probíhá na základě elektrotonického šíření elektrického potenciálu. Když presynaptický neuron vygeneruje akční potenciál nebo i jen podprahovou depolarizaci, vznikne rozdíl elektrického potenciálu mezi oběma stranami spojení. Ionty pak přirozeně proudí z místa vyššího potenciálu do místa nižšího, a to přímo skrze gap junction kanálky. Výsledkem je, že postsynaptický neuron zaznamená téměř okamžitou změnu svého membránového potenciálu. Tato rychlost je klíčovou výhodou elektrických synapsí, protože přenos probíhá prakticky bez zpoždění, na rozdíl od chemických synapsí, kde synaptická prodleva trvá alespoň několik desetin milisekundy.
Důležitou vlastností elektrických synapsí je jejich obousměrnost. Zatímco chemické synapsy přenášejí signál vždy jen jedním směrem, od presynaptického zakončení k postsynaptickému receptoru, elektrické synapsy umožňují tok iontů oběma směry. To znamená, že pokud se depolarizuje postsynaptický neuron, ovlivní to i neuron presynaptický. Tato vlastnost má zásadní důsledky pro koordinaci neuronálních sítí a synchronizaci aktivity skupin neuronů. Existují sice i rektifikující elektrické synapsy, které preferují přenos v jednom směru díky asymetrickému složení konexinů, ale obecně platí, že bidirektionalita je pro elektrické synapsy charakteristická.
V kontextu neuronálních sítí hrají elektrické synapsy nezastupitelnou roli při synchronizaci skupin neuronů. Například v mozečku, hipokampu nebo v čichových bulbech jsou elektrické synapsy hojně zastoupeny mezi interneurony, kde zajišťují koordinovanou aktivitu celých neuronálních populací. Tato synchronizace je základem pro vznik oscilačních rytmů, jako jsou gama oscilace, které jsou spojovány s kognitivními procesy, pozorností a vědomím. Bez elektrických synapsí by taková přesná časová koordinace byla jen stěží dosažitelná.
Elektrické synapsy nejsou statickými strukturami. Jejich propustnost a funkční stav jsou dynamicky regulovány celou řadou faktorů. Intracelulární pH, koncentrace vápenatých iontů, fosforylace konexinových proteinů prostřednictvím různých kináz nebo přítomnost specifických ligandů mohou výrazně ovlivnit, jak dobře kanálky vedou elektrický proud. To znamená, že elektrické synapsy nejsou pouhými pasivními vodivými spoji, ale aktivními prvky nervového systému, schopnými přizpůsobovat svou funkci aktuálním potřebám buňky a celé sítě.
Zajímavým aspektem je také vývojová regulace elektrických synapsí. V raném embryonálním vývoji jsou gap junctions mezi neurony velmi hojné a hrají klíčovou roli při koordinaci buněčného dělení, migraci neuronů a tvorbě základní architektury nervového systému. S postupující maturací mozku se část elektrických synapsí nahrazuje synapsemi chemickými, avšak mnohé zůstávají zachovány a plní specifické funkce v dospělém organismu. Mutace v genech kódujících konexiny jsou spojeny s celou řadou neurologických onemocnění, včetně epilepsie nebo periferních neuropatií, což podtrhuje jejich fyziologický význam.
Přímý přenos iontů elektrickými synapsemi tak představuje elegantní a evolučně konzervovaný mechanismus, který doplňuje a v mnoha ohledech obohacuje možnosti nervové soustavy. Jeho pochopení je nezbytné pro komplexní vnímání toho, jak neurony komunikují, jak se synchronizují a jak společně vytvářejí složité vzorce aktivity, jež stojí za veškerým vnímáním, myšlením a chováním živých organismů.
Synaptické váčky představují jednu z nejzásadnějších struktur celého nervového systému, přičemž jejich existence a funkčnost jsou naprosto nepostradatelné pro správné přenášení informací mezi neurony. Tyto drobné membránové struktury, které se nacházejí v presynaptickém zakončení nervové buňky, v sobě ukrývají neurotransmitery – chemické látky, jež slouží jako poslové mezi jednotlivými nervovými buňkami. Bez jejich přítomnosti by komunikace v nervové soustavě jednoduše nemohla fungovat tak, jak ji známe.
Každý neuron je schopen přijímat a odesílat signály prostřednictvím synapsí, a právě v tomto procesu hrají synaptické váčky zcela klíčovou roli. Když nervový impuls dorazí do presynaptického zakončení, spustí se kaskáda dějů, která vede k uvolnění obsahu těchto váčků do synaptické štěrbiny. Tento proces se nazývá exocytóza a je nesmírně přesně regulován. Váčky se přiblíží k presynaptické membráně, splyne s ní a neurotransmitery jsou uvolněny do prostoru mezi dvěma neurony – do oné úzké synaptické štěrbiny, která měří pouhých několik desítek nanometrů.
Synaptické váčky nejsou náhodně rozmístěné struktury – jsou organizovány do specifických zón, které se nazývají aktivní zóny. Právě v těchto místech dochází k největší pravděpodobnosti fúze váčku s membránou a k následnému uvolnění neurotransmiterů. Tato organizace zajišťuje, že přenos signálu je rychlý, efektivní a prostorově přesný.
Po uvolnění neurotransmiterů do synaptické štěrbiny se tyto látky váží na receptory postsynaptické membrány sousedního neuronu. Tato vazba způsobuje změny v propustnosti iontových kanálů postsynaptické buňky, což vede buď k excitaci, nebo inhibici tohoto neuronu – záleží na tom, jaký typ neurotransmiteru byl uvolněn a jaký receptor se na postsynaptické membráně nachází. Celý tento proces probíhá v řádu milisekund a umožňuje nervové soustavě reagovat na podněty s ohromující rychlostí.
Co je na synaptických váčcích fascinující, je jejich schopnost recyklace. Po uvolnění svého obsahu nejsou váčky jednoduše zničeny – jejich membrána je znovu začleněna do presynaptické membrány a následně recyklována prostřednictvím procesu zvaného endocytóza. Tímto způsobem si neuron udržuje dostatečnou zásobu váčků připravených k dalšímu použití, aniž by musel neustále syntetizovat nové membránové komponenty od základu. Tento elegantní mechanismus recyklace je jedním z příkladů toho, jak evoluce optimalizovala fungování nervového systému.
Neurotransmitery, které jsou v synaptických váčcích uloženy, jsou velmi různorodé. Patří mezi ně například glutamát, který je hlavním excitačním neurotransmiterem v centrálním nervovém systému, dále kyselina gama-aminomáselná neboli GABA, jež naopak působí inhibičně, a také acetylcholin, dopamin, serotonin či noradrenalin. Každý z těchto neurotransmiterů je uložen ve specifickém typu váčků a plní odlišné funkce v rámci nervové soustavy.
Poruchy ve fungování synaptických váčků mohou mít závažné důsledky pro zdraví člověka. Celá řada neurologických a psychiatrických onemocnění je spojena s dysfunkcí synaptického přenosu. Například Parkinsonova choroba je charakterizována nedostatkem dopaminu v určitých oblastech mozku, deprese bývá spojována s nerovnováhou serotoninu a noradrenalinu, a schizofrenické příznaky jsou dávány do souvislosti s dysregulací dopaminergního systému. Mnohé léky, které se používají k léčbě těchto stavů, působí právě na úrovni synaptického přenosu – ovlivňují uvolňování neurotransmiterů, jejich zpětné vstřebávání nebo blokují příslušné receptory.
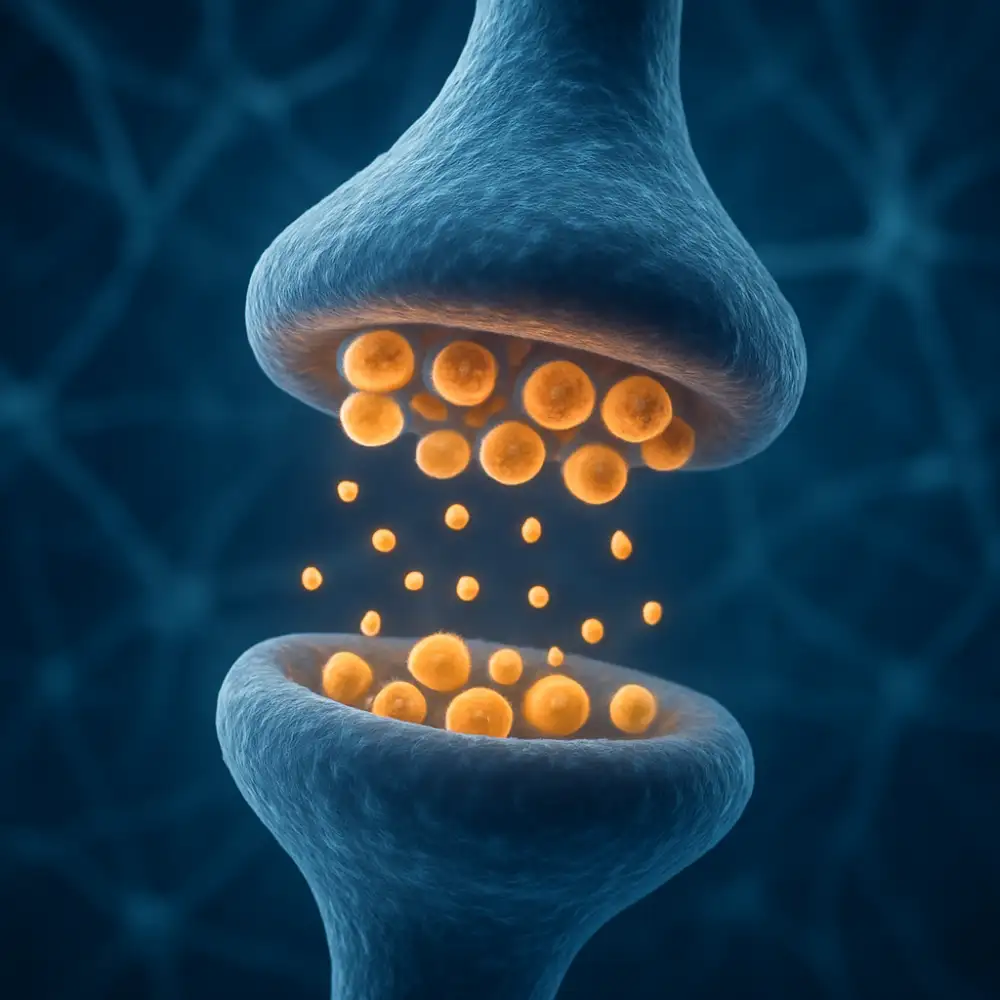
Je také důležité zmínit, že synaptická plasticita – tedy schopnost synapsí měnit svou sílu a efektivitu v závislosti na předchozí aktivitě – je úzce spjata s funkcí synaptických váčků. Dlouhodobá potenciace, která je považována za jeden ze základních mechanismů učení a paměti, zahrnuje změny v počtu a rozmístění synaptických váčků, v citlivosti postsynaptických receptorů i v celkové architektuře synapse. Synaptické váčky jsou tedy nejen přenašeči okamžitých signálů, ale také klíčovými hráči v procesech, které formují naši schopnost učit se a pamatovat si.
Postsynaptická membrána představuje vysoce specializovanou strukturu, která je klíčovým prvkem v procesu synaptického přenosu. Její funkce závisí na přítomnosti specifických receptorů, jež jsou schopny rozpoznat a vázat konkrétní neurotransmitery uvolňované z presynaptického zakončení. Tato specifičnost není náhodná – je výsledkem milionů let evoluce a představuje jeden z nejsofistikovanějších biochemických systémů v celém živém světě.
Receptory postsynaptické membrány jsou proteinové komplexy zabudované přímo do lipidové dvojvrstvy membrány, přičemž jejich prostorová konfigurace umožňuje selektivní vazbu pouze určitých molekul. Každý receptor má takzvanou vazebnou doménu, která svým tvarem přesně odpovídá struktuře příslušného neurotransmiteru – hovoříme o principu klíče a zámku. Jakmile se neurotransmiter naváže na receptor, dochází ke konformační změně proteinu, která spouští kaskádu dalších dějů uvnitř postsynaptické buňky.
Z hlediska funkčního dělení rozlišujeme dva základní typy postsynaptických receptorů. Ionotropní receptory fungují jako ligand-řízené iontové kanály, což znamená, že jejich aktivace vede k okamžitému otevření kanálu a průtoku iontů přes membránu. Tento proces je velmi rychlý a probíhá v řádu milisekund. Typickým příkladem jsou receptory pro glutamát, konkrétně receptory AMPA a NMDA, nebo receptory pro kyselinu gama-aminomáselnou, tedy GABA-A receptory. Metabotropní receptory naproti tomu působí prostřednictvím druhých poslů a jejich efekt nastupuje pomaleji, ale trvá déle. Tyto receptory jsou spřaženy s G-proteiny, které po aktivaci spouštějí intracelulární signální dráhy vedoucí k různým metabolickým změnám.
Specifičnost receptorů má zásadní význam pro správnou funkci neuronálních sítí. Jeden neuron může přijímat signály od tisíců dalších neuronů, přičemž každý z nich může uvolňovat odlišný neurotransmiter. Postsynaptická membrána musí být schopna tyto signály rozlišit a adekvátně na ně reagovat. Proto je rozmístění různých typů receptorů na membráně přísně regulováno a není rovnoměrné – v oblasti postsynaptické denzity se koncentrují specifické receptory odpovídající neurotransmiterům, které daná synapse využívá.
Důležitou roli hrají také pomocné proteiny, jako jsou scaffoldingové proteiny, které ukotvují receptory na správném místě a udržují jejich správnou hustotu. PSD-95 je jedním z nejlépe prostudovaných scaffoldingových proteinů, který se váže na NMDA receptory a zajišťuje jejich stabilní pozici v postsynaptické denzitě. Bez těchto pomocných molekul by receptory difundovaly po membráně a synaptický přenos by byl výrazně méně efektivní.
Plasticita synaptických receptorů je základem učení a paměti. Při opakované aktivaci synapse může docházet ke zvýšení počtu receptorů na postsynaptické membráně, což vede k posílení synaptického spojení – tento jev nazýváme dlouhodobou potenciací. Opačný proces, dlouhodobá deprese, zahrnuje snížení počtu nebo citlivosti receptorů. Oba tyto mechanismy jsou závislé na přesné regulaci receptorové specifičnosti a jejich dynamickém přeskupování v membráně.
Poruchy v expresi nebo funkci postsynaptických receptorů jsou spojeny s celou řadou neurologických a psychiatrických onemocnění. Schizofrenie, deprese, epilepsie nebo Alzheimerova choroba – všechna tato onemocnění mají alespoň částečně svůj základ v dysfunkci synaptických receptorů. Proto je pochopení jejich specifičnosti a mechanismů regulace nejen akademicky zajímavé, ale má i přímý klinický dopad na vývoj nových terapeutických přístupů.
Každý neuron v nervovém systému je neustále bombardován obrovským množstvím signálů přicházejících z tisíců dalších neuronů, přičemž tyto signály mohou mít buď povzbuzující, nebo tlumivý charakter. Právě tato dualita tvoří základ veškeré nervové komunikace a umožňuje mozku zpracovávat informace s neobyčejnou přesností a flexibilitou. Synaptické potenciály představují základní funkční jednotku této komunikace a jejich pochopení je klíčové pro porozumění tomu, jak nervová soustava jako celek funguje.
Když presynaptický neuron vyšle akční potenciál, dojde k uvolnění neurotransmiterů do synaptické štěrbiny. Tyto chemické posly se následně vážou na receptory postsynaptické membrány, čímž spouštějí kaskádu dějů, které vedou ke změně elektrického potenciálu postsynaptické buňky. Výsledkem tohoto procesu může být buď excitační postsynaptický potenciál (EPSP), nebo inhibiční postsynaptický potenciál (IPSP), přičemž každý z těchto typů má zcela odlišný vliv na pravděpodobnost vzniku nového akčního potenciálu v postsynaptickém neuronu.
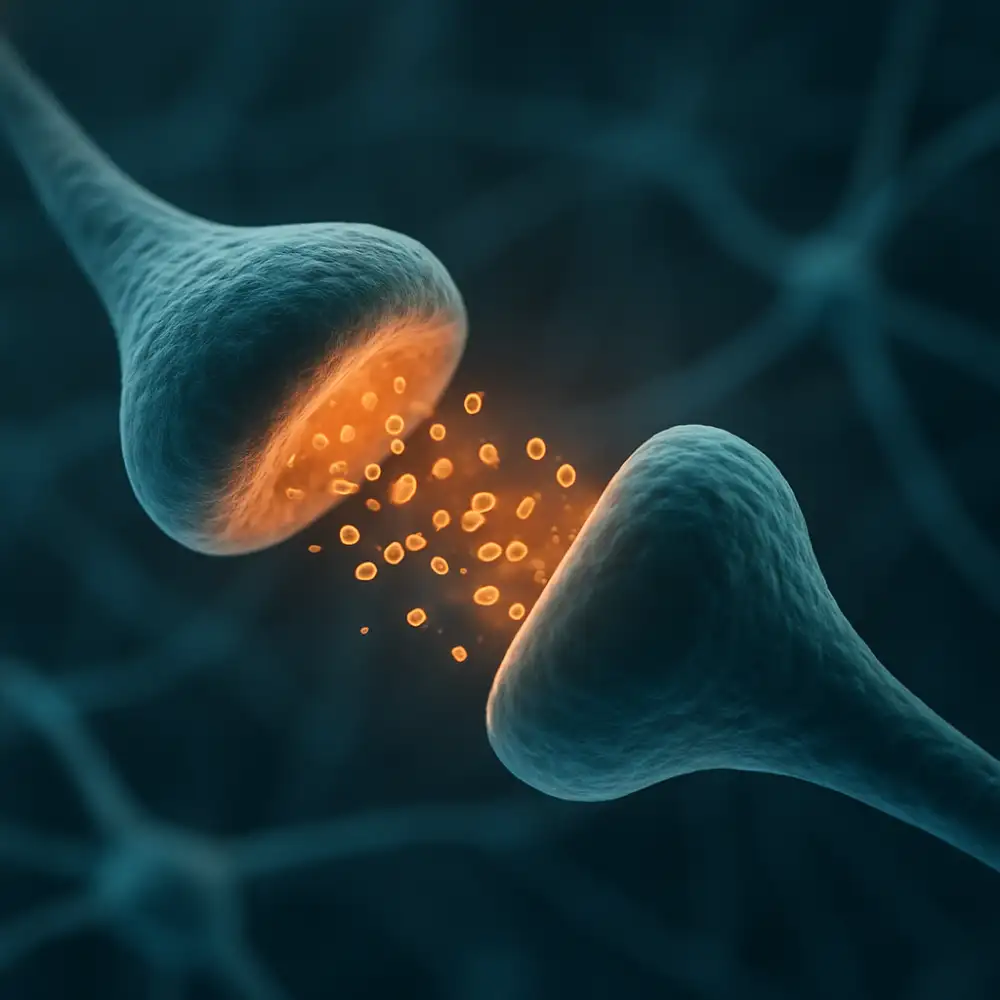
Excitační postsynaptický potenciál vzniká tehdy, když neurotransmiter způsobí depolarizaci postsynaptické membrány, tedy přiblížení membránového potenciálu k prahovým hodnotám potřebným pro vznik akčního potenciálu. Nejčastěji k tomu dochází otevřením iontových kanálů propustných pro sodíkové ionty, které proudí do buňky po svém koncentračním gradientu a způsobují tak lokální kladnou změnu membránového potenciálu. Glutamát je v tomto kontextu nejvýznamnějším excitačním neurotransmiterem v centrálním nervovém systému a jeho působení prostřednictvím AMPA a NMDA receptorů je předmětem intenzivního výzkumu zejména v souvislosti s mechanismy učení a paměti.
Naproti tomu inhibiční postsynaptický potenciál vzniká hyperpolarizací postsynaptické membrány, tedy jejím vzdálením od prahových hodnot, což snižuje pravděpodobnost vzniku akčního potenciálu. Tento efekt je nejčastěji zprostředkován otevřením chloridových kanálů, přičemž chloridové ionty proudí do buňky a způsobují zápornou změnu membránového potenciálu. Kyselina gama-aminomáselná, obecně známá jako GABA, je hlavním inhibičním neurotransmiterem mozku a hraje zásadní roli v regulaci neuronální excitability. Narušení rovnováhy mezi excitačními a inhibičními signály může vést k závažným neurologickým poruchám, včetně epilepsie nebo úzkostných stavů.
Zásadní aspekt synaptické komunikace spočívá v tom, že jednotlivý neuron integruje všechny příchozí signály v procesu nazývaném sumace. Rozlišujeme přitom dva základní typy sumace. Temporální sumace nastává tehdy, když jeden presynaptický neuron vyšle rychle za sebou více signálů, jejichž účinky se na postsynaptické membráně sčítají dříve, než dojde k jejich odeznění. Prostorová sumace naproti tomu zahrnuje současné působení více různých presynaptických neuronů na tentýž postsynaptický neuron, přičemž výsledný efekt závisí na poměru excitačních a inhibičních vstupů.
Axonový hrbolek, neboli axon hillock, je místem, kde dochází k finálnímu rozhodnutí o tom, zda neuron vyšle akční potenciál, či nikoli. Právě zde se sčítají všechny postsynaptické potenciály a pokud jejich výsledný součet překročí prahovou hodnotu přibližně minus padesát pět milivoltů, vzniká akční potenciál, který se šíří dále po axonu k dalším synapsím. Tato integrace je nesmírně sofistikovaný proces, neboť neuron musí neustále vyvažovat stovky až tisíce excitačních a inhibičních vstupů přicházejících v různých časových okamžicích a z různých částí jeho dendritického stromu.
Modulace synaptické přenosu hraje rovněž klíčovou roli v plasticitě nervového systému. Dlouhodobá potenciace, označovaná zkratkou LTP, představuje proces, při němž opakovaná aktivace synapse vede k trvalému zvýšení její účinnosti, zatímco dlouhodobá deprese, LTD, má opačný efekt. Tyto jevy jsou považovány za buněčný základ učení a paměti a jsou úzce spjaty s dynamikou excitačních a inhibičních synaptických potenciálů. Například NMDA receptory, které jsou propustné pro vápenatý ionty, hrají klíčovou roli v indukci LTP, neboť fungují jako molekulární koincidencní detektory schopné zaregistrovat souběžnou aktivitu presynaptického i postsynaptického neuronu.
Rovnováha mezi excitací a inhibicí není statická, ale neustále se dynamicky přizpůsobuje aktuálním potřebám nervového systému. Tato rovnováha je regulována na mnoha úrovních, od rychlých změn v propustnosti iontových kanálů až po pomalejší modulační mechanismy zprostředkované metabotropními receptory a druhými posly. Pochopení těchto mechanismů má zásadní klinický význam, neboť mnohé psychofarmaka, anestetika i drogy působí právě prostřednictvím ovlivnění excitačních nebo inhibičních synaptických přenosů, čímž mění celkový stav vědomí, náladu nebo vnímání bolesti.
Každý synaptický spoj mezi neurony je jako tichý most, po němž přechází nejen elektrický impuls, ale i samotná podstata našeho myšlení, cítění a vědomí – bez těchto nepatrných spojení by mozek byl pouhým tichým ostrovem uprostřed moře nevyužitého potenciálu.
Rostislav Dvořáček
Mozek je jedním z nejsložitějších systémů, jaké kdy příroda vytvořila, a jeho schopnost učit se a přizpůsobovat okolnímu světu spočívá z velké části právě v procesech, které se odehrávají na úrovni synapsí. Každý neuron v lidském mozku je spojen s tisíci dalšími neurony prostřednictvím specializovaných spojů, které nazýváme synapsemi, a právě tyto spoje jsou místem, kde se rodí paměť, učení i veškeré kognitivní schopnosti člověka.
Synaptická plasticita je schopnost synapsí měnit svou sílu a efektivitu v závislosti na předchozí aktivitě. Tento jev byl poprvé systematicky popsán v kontextu Hebbova pravidla, které říká, že neurony, jež se aktivují společně, posilují svá vzájemná spojení. Jinými slovy, pokud jeden neuron opakovaně přispívá k aktivaci druhého neuronu, synaptické spojení mezi nimi se postupně zesiluje. Tento princip je základním kamenem toho, jak mozek ukládá informace a jak se učí novým dovednostem.
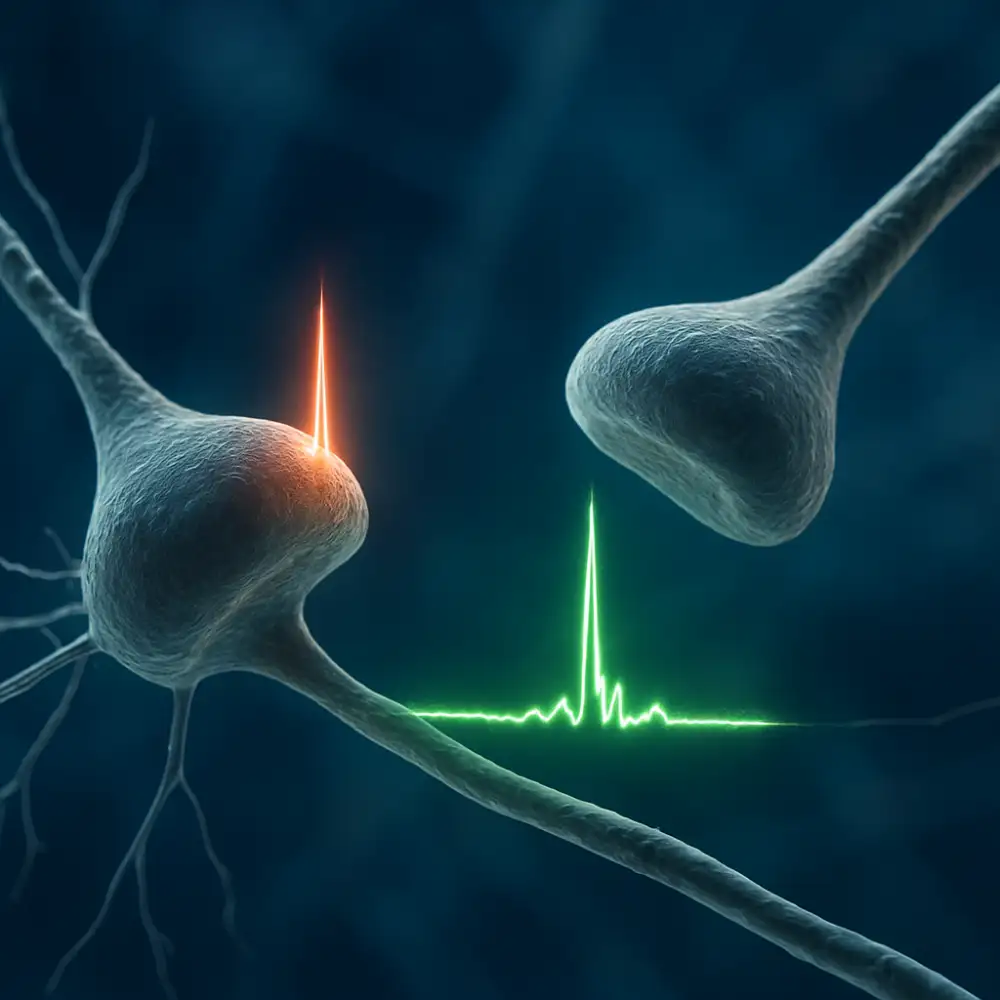
Na molekulární úrovni se synaptická plasticita projevuje především prostřednictvím dvou klíčových procesů – dlouhodobé potenciace (LTP) a dlouhodobé deprese (LTD). Dlouhodobá potenciace představuje trvalé zesílení synaptického přenosu, ke kterému dochází po opakované nebo intenzivní stimulaci. Klíčovou roli v tomto procesu hrají NMDA receptory, které fungují jako jakési molekulární detektory koincidence – aktivují se pouze tehdy, když jsou presynaptický i postsynaptický neuron aktivní současně. Jakmile jsou tyto receptory aktivovány, spouštějí kaskádu biochemických reakcí, které vedou k zabudování dalších AMPA receptorů do postsynaptické membrány, čímž se zvyšuje citlivost synapsy na budoucí signály.
Proces přenosu signálu ze synapse na synapse je fascinujícím příkladem biologické přesnosti a komplexity. Když akční potenciál dorazí do presynaptického zakončení, způsobí otevření napěťově řízených vápníkových kanálů. Vápník, který proudí dovnitř buňky, spouští fúzi synaptických váčků s presynaptickou membránou, čímž dochází k uvolnění neurotransmiterů do synaptické štěrbiny. Tyto molekuly pak difundují přes štěrbinu a váží se na receptory postsynaptického neuronu, kde vyvolávají elektrické nebo biochemické změny.
Synaptická plasticita se neomezuje pouze na zesílení spojení. Stejně důležitá je i schopnost mozku oslabovat nebo dokonce eliminovat synaptická spojení, která nejsou využívána. Tento proces, označovaný jako synaptická deprese nebo synaptické prořezávání, je nezbytný pro efektivní fungování nervové sítě. Mozek, který by si udržoval všechna synaptická spojení bez ohledu na jejich relevanci, by byl zahlcen nadbytečnými informacemi a nebyl by schopen efektivně zpracovávat nové podněty.
V průběhu vývoje mozku, zejména v dětském věku, probíhá intenzivní synaptogeneze – tvorba nových synaptických spojení. Dítě se rodí s relativně malým počtem synapsí, ale v průběhu prvních let života jejich počet dramaticky narůstá. Tento proces je řízen kombinací genetických faktorů a zkušeností z okolního prostředí, přičemž senzitivní periody vývoje představují okna, během nichž je mozek obzvláště vnímavý na určité typy podnětů. Po skončení těchto senzitivních period dochází k selektivnímu prořezávání synapsí – přežívají ta spojení, která jsou opakovaně aktivována, zatímco nepoužívaná spojení zanikají.
Hippokampus hraje v procesech synaptické plasticity a učení naprosto klíčovou roli. Tato oblast mozku, která svým tvarem připomíná mořského koníka, je nezbytná pro tvorbu nových deklarativních pamětí. Právě v hippokampu byla poprvé experimentálně prokázána dlouhodobá potenciace, a právě zde probíhají některé z nejintenzivnějších synaptických přeměn při učení. Neurony hippokampu jsou schopny velmi rychle reagovat na nové informace a vytvářet nové synaptické spoje v průběhu hodin nebo dokonce minut.
Zajímavým aspektem synaptické plasticity je její závislost na časování. Spike-timing dependent plasticity, neboli plasticita závislá na časování akčních potenciálů, ukazuje, že záleží nejen na tom, zda se dva neurony aktivují společně, ale také na tom, v jakém pořadí se aktivují. Pokud presynaptický neuron vyšle signál těsně před tím, než se aktivuje postsynaptický neuron, synaptické spojení se posiluje. Pokud je pořadí opačné, synaptické spojení se oslabuje. Tento mechanismus umožňuje mozku kódovat nejen asociace mezi podněty, ale také jejich časové vztahy a kauzální strukturu.
Moderní neurověda stále odkrývá nové vrstvy složitosti synaptické plasticity. Bylo zjištěno, že synaptická plasticita není jen záležitostí jednotlivých synapsí, ale že probíhá na úrovni celých neuronálních sítí, kde se vzájemně ovlivňují tisíce synaptických spojení najednou. Homestatická plasticita, což je schopnost neuronu regulovat svou celkovou aktivitu tak, aby zůstala v optimálním rozsahu, představuje další vrstvu regulace, která zajišťuje stabilitu nervové sítě i při intenzivním učení. Bez těchto homeostatických mechanismů by nadměrná potenciace mohla vést k epileptické aktivitě nebo naopak k úplnému utlumení neuronální aktivity.
Mozek je orgán, který nikdy nepřestává překvapovat svou schopností přizpůsobovat se a uchovávat informace. Jedním z nejzásadnějších mechanismů, které stojí za naší schopností učit se a pamatovat si, je proces označovaný jako dlouhodobá potenciace, zkráceně LTP z anglického long-term potentiation. Tento fenomén byl poprvé popsán v sedmdesátých letech dvacátého století norským neurovědcem Terje Lømo a od té doby se stal jedním z nejvíce studovaných procesů v celé neurobiologii.
Abychom pochopili, jak dlouhodobá potenciace funguje, musíme se nejprve zaměřit na základní stavební jednotku nervového systému, tedy na neuron a jeho synapse. Synapse je místo, kde dochází k přenosu informací mezi dvěma neurony. Presynaptický neuron vysílá signál ve formě akčního potenciálu, který způsobí uvolnění neurotransmiterů do synaptické štěrbiny. Tyto chemické posly pak putují přes štěrbinu a váží se na receptory postsynaptického neuronu, čímž v něm vyvolávají odpovídající elektrický signál. Tento zdánlivě jednoduchý proces je ve skutečnosti nesmírně komplexní a jeho modifikace leží v samém srdci toho, co nazýváme pamětí.
Klíčovým hráčem v procesu dlouhodobé potenciace je receptor NMDA, neboli N-methyl-D-aspartátový receptor. Tento receptor je velmi specifický v tom, že pro svou aktivaci vyžaduje splnění dvou podmínek současně. Jednak musí být přítomen glutamát, který se váže na receptor, a jednak musí být postsynaptická membrána dostatečně depolarizována, aby byl uvolněn hořečnatý iont, který za klidových podmínek blokuje iontový kanál receptoru. Tato vlastnost z receptoru NMDA dělá jakýsi molekulární detektor koincidence, který reaguje pouze tehdy, když jsou presynaptický a postsynaptický neuron aktivní ve stejnou dobu. Právě tato vlastnost je základem Hebbova pravidla, které říká, že neurony, které se aktivují společně, se propojují dohromady.
Když jsou tedy oba neurony aktivní současně a receptor NMDA se otevře, dochází k masivnímu přílivu vápenatých iontů do postsynaptické buňky. Vápenatý iont funguje jako druhý posel a spouští celou kaskádu intracelulárních signálních drah. Jednou z nejdůležitějších je aktivace enzymu CaMKII, kalcium-kalmodulin dependentní proteinkinázy II. Tento enzym fosforyluje receptory AMPA, čímž zvyšuje jejich vodivost, a navíc podporuje jejich přesun do synaptické membrány. Výsledkem je, že postsynaptický neuron reaguje na stejné množství glutamátu silněji než před stimulací. Synapse se stala potencovanou.
Dlouhodobá potenciace však není jen krátkodobou záležitostí. Pokud je stimulace dostatečně silná nebo opakovaná, dochází k aktivaci genové exprese a syntéze nových proteinů, které vedou ke strukturálním změnám synapse. Dendritické trny, malé výběžky na dendritech postsynaptického neuronu, se zvětšují a mění svůj tvar. Mohou se dokonce vytvářet zcela nové synaptické spoje. Tato strukturální plasticita představuje fyzický substrát dlouhodobé paměti a je důvodem, proč si dokážeme pamatovat události z dávné minulosti.
Fascinující je také to, jak synapse komunikuje zpětně s presynaptickým neuronem. Postsynaptický neuron totiž nevystupuje jen jako pasivní příjemce informací. V průběhu dlouhodobé potenciace produkuje retrográdní posly, jako je oxid dusnatý nebo endokanabinoidy, které putují zpět přes synaptickou štěrbinu a ovlivňují presynaptický terminál. Tímto způsobem dochází k posílení celého synaptického spojení z obou stran, přičemž presynaptický neuron začne uvolňovat více neurotransmiterů při každé aktivaci.
Je důležité si uvědomit, že paměť není uložena v jednom neuronu, ale v celých sítích neuronů propojených synapsemi. Každá vzpomínka odpovídá specifickému vzorci aktivace neuronální sítě, přičemž síla jednotlivých synaptických spojení v této síti určuje, jak snadno a přesně si danou vzpomínku dokážeme vybavit. Opakovaná aktivace určité sítě prostřednictvím učení nebo vzpomínání posiluje příslušné synaptické spoje a usnadňuje budoucí aktivaci téže sítě. To je důvod, proč opakování látky zlepšuje zapamatování.
Výzkum dlouhodobé potenciace také odhalil, proč jsou emocionálně nabité vzpomínky tak trvalé a živé. Amygdala, mozková struktura zodpovědná za zpracování emocí, dokáže modulovat sílu synaptické plasticity v hippokampu, který je klíčový pro tvorbu explicitních vzpomínek. Stresové hormony jako adrenalin a kortizol zvyšují pravděpodobnost indukce dlouhodobé potenciace, čímž zajišťují, že emocionálně důležité události jsou uloženy přednostně a s větší intenzitou.
Pochopení mechanismů dlouhodobé potenciace má obrovský praktický význam. Poruchy synaptické plasticity jsou spojovány s celou řadou neurologických a psychiatrických onemocnění, včetně Alzheimerovy choroby, schizofrenie nebo deprese. Výzkumníci po celém světě se snaží vyvinout farmakologické přístupy, které by dokázaly cíleně modulovat procesy synaptické plasticity a tím pomoci pacientům s poruchami paměti nebo učení. Každý nový objev v oblasti molekulárních mechanismů LTP tak otevírá nové terapeutické možnosti a přibližuje nás k hlubšímu pochopení toho, co nás dělá tím, čím jsme.
Synaptický přenos představuje jeden z nejkomplexnějších a zároveň nejzranitelnějších procesů v celém nervovém systému. Jakákoli porucha v přenosu signálu mezi neurony může mít dalekosáhlé důsledky pro fungování mozku i celého těla. Nervová soustava je závislá na precizní koordinaci synaptické komunikace, a proto i zdánlivě drobné odchylky v tomto procesu mohou vést k závažným neurologickým nebo psychiatrickým onemocněním.
Jedním z nejlépe prostudovaných příkladů je Parkinsonova choroba, při níž dochází k postupnému úbytku dopaminergních neuronů v oblasti substantia nigra. Tyto neurony jsou zodpovědné za uvolňování dopaminu do striata, přičemž dopamin hraje klíčovou roli v regulaci pohybů. Když synaptický přenos dopaminu selže nebo je výrazně omezen, projevuje se to třesem, svalovou rigiditou a bradykinezí. Synapse mezi dopaminergními neurony a neurony striata přestávají fungovat správně, což narušuje celý motorický okruh bazálních ganglií. Léčba levodopou se snaží tento deficit kompenzovat, avšak s postupující degenerací neuronů se stává méně účinnou.
Podobně devastující dopady má Alzheimerova choroba, která je charakterizována nejen tvorbou amyloidových plaků a neurofibrilárních klubek, ale také výraznou ztrátou synaptických spojení. Výzkumy opakovaně ukazují, že ztráta synapsí koreluje s kognitivním úpadkem lépe než samotná přítomnost plaků. Cholinergní neurony bazálního předního mozku, které zásobují hippokampus a mozkovou kůru acetylcholinem, jsou postiženy jako jedny z prvních. Narušení synaptického přenosu acetylcholinu vede k poruchám paměti a učení, protože hippokampální synapse jsou nezbytné pro konsolidaci paměťových stop.
Epilepsie představuje další skupinu onemocnění, kde je synaptická dysbalance zcela zásadní. Při epileptickém záchvatu dochází k nekontrolovanému, synchronnímu výboji velkého počtu neuronů. Přílišná excitace zprostředkovaná glutamátergními synapsemi nebo nedostatečná inhibice zprostředkovaná GABAergními synapsemi může spustit tento patologický stav. Mutace v genech kódujících iontové kanály nebo receptory na postsynaptické membráně jsou u mnoha forem epilepsie přímo odpovědné za vzniklou dysbalanci. Například mutace v genech pro GABA-A receptor nebo pro napěťově řízené sodíkové kanály mění práh pro vznik akčního potenciálu a narušují normální synaptický přenos.
Roztroušená skleróza sice primárně postihuje myelinové pochvy axonů, avšak sekundárně výrazně ovlivňuje i synaptický přenos. Demyelinizace zpomaluje nebo zcela blokuje vedení nervového vzruchu, takže signál buď nedorazí k synaptickému zakončení vůbec, nebo dorazí se zpožděním a v pozměněné podobě. Synapse, které jsou na konci takto poškozených axonů, pak nemohou plnit svou funkci, a to se projevuje celou škálou neurologických příznaků od poruchy zraku přes svalovou slabost až po kognitivní problémy.
Schizofrenie a další psychiatrická onemocnění jsou rovněž úzce spjata s poruchami synaptického přenosu, i když mechanismy jsou zde složitější a méně jednoznačné. Hypotéza dopaminové dysregulace předpokládá, že nadměrná dopaminergní aktivita v mezolimbickém systému přispívá k pozitivním příznakům schizofrenie, zatímco snížená dopaminergní aktivita v prefrontální kůře je odpovědná za negativní příznaky a kognitivní deficit. Glutamátergní hypotéza pak zdůrazňuje roli NMDA receptorů, jejichž hypofunkce narušuje normální synaptický přenos a vede k podobným příznakům jako schizofrenie.
Myasthenia gravis je autoimunitní onemocnění, při němž protilátky napadají nikotinové acetylcholinové receptory na nervosvalové ploténce. Tato specializovaná synapse mezi motorickým neuronem a svalovou buňkou je tak poškozena, že přenos signálu selhává a sval se nedokáže dostatečně stáhnout. Výsledkem je svalová slabost, která se typicky zhoršuje při opakované aktivitě, protože zásoby acetylcholinu v presynaptickém zakončení se postupně vyčerpávají a jejich doplňování nestačí kompenzovat blokádu receptorů.
Výzkum synaptického přenosu a jeho poruch přináší stále nové poznatky, které otevírají cestu k vývoji cílených terapií. Pochopení molekulárních mechanismů synaptické dysfunkce je klíčem k léčbě celé řady neurologických a psychiatrických onemocnění, která dosud zůstávají jen částečně zvladatelná. Každý neuron, každá synapse a každý přenašeč hrají svou nezastupitelnou roli v obrovské síti nervového systému, a narušení byť jediného článku tohoto řetězce může mít pro pacienta zásadní důsledky.
Věda o synapsích prošla v posledních desetiletích dramatickým vývojem, který zásadně proměnil naše chápání toho, jak neurony mezi sebou komunikují a jak tato komunikace ovlivňuje zdraví i nemoc. Dnes víme, že synapse není pouhým místem přenosu elektrického signálu, ale představuje složitý dynamický systém, který se neustále přizpůsobuje, mění a reaguje na podněty z okolního prostředí. Synaptická plasticita, tedy schopnost synapsí měnit svou sílu a efektivitu v závislosti na předchozí aktivitě, stojí v centru moderního neurovědeckého výzkumu a otevírá zcela nové obzory v oblasti léčby neurologických a psychiatrických onemocnění.
Jedním z klíčových průlomů posledních let je podrobné mapování takzvaného synaptického proteómu, tedy souboru všech proteinů, které se nacházejí v synaptické štěrbině a v postsynaptické hustotě. Vědci z různých světových pracovišť zjistili, že v jediné synapse může být přítomno více než tisíc různých proteinů, přičemž každý z nich plní specifickou funkci při regulaci přenosu signálu. Poruchy v expresi nebo funkci těchto proteinů jsou dnes spojovány s celou řadou závažných onemocnění, včetně schizofrenie, autismu, Alzheimerovy choroby nebo deprese. Tento poznatek otevřel zcela nové terapeutické cíle, na které mohou být zaměřena nová farmaka.
Výzkum glutamátergních synapsí, tedy synapsí využívajících jako neurotransmiter glutamát, přinesl zvláště pozoruhodné výsledky. Receptory NMDA a AMPA, které jsou klíčovými mediátory excitační synaptické transmise, se ukázaly být zásadní nejen pro normální kognitivní funkce, ale také pro patofyziologii celé řady duševních poruch. Ketamin, látka původně používaná jako anestetikum, prokázal překvapivě rychlé antidepresivní účinky právě skrze svůj vliv na NMDA receptory, což vedlo k vývoji nové generace antidepresiv, jejichž mechanismus účinku se zásadně liší od klasických selektivních inhibitorů zpětného vychytávání serotoninu.
Paralelně s tím probíhá intenzivní výzkum GABAergních synapsí, které zprostředkovávají inhibiční přenos v centrálním nervovém systému. Narušení rovnováhy mezi excitačními a inhibičními synapsemi, označované jako E/I imbalance, je považováno za jeden ze základních neurobiologických mechanismů, které stojí v pozadí poruch autistického spektra nebo epilepsie. Cílená modulace GABAergní transmise pomocí nových farmakologických nástrojů představuje slibnou cestu k efektivnější léčbě těchto stavů, aniž by docházelo k nežádoucím vedlejším účinkům spojeným se staršími léčivy.
Moderní zobrazovací techniky, jako je super-rozlišovací mikroskopie nebo dvoufotonová mikroskopie in vivo, umožnily vědcům pozorovat synaptické procesy v reálném čase a s dosud nevídaným prostorovým rozlišením. Díky těmto metodám bylo možné přímo sledovat, jak se synaptické váčky pohybují k presynaptické membráně, jak probíhá exocytóza neurotransmiterů a jak postsynaptická hustota reorganizuje své složení v průběhu synaptické plasticity. Tyto poznatky jsou naprosto zásadní pro pochopení toho, co přesně se děje na synaptické úrovni při učení a paměti, ale také při neurodegenerativních procesech.
V kontextu Alzheimerovy choroby je obzvláště důležité zjištění, že synaptická dysfunkce předchází samotné ztrátě neuronů o mnoho let a že právě narušení synaptické transmise koreluje nejlépe s kognitivním deficitem pacientů. Beta-amyloidní oligomery, které se hromadí v mozku nemocných, se vážou přímo na synaptické proteiny a narušují funkci NMDA receptorů, čímž blokují normální mechanismy synaptické plasticity. Terapeutické strategie zaměřené na ochranu synapsí před tímto poškozením jsou proto považovány za perspektivnější než pouhé snižování celkového amyloidního zatížení mozku.
Výzkum synapsí dnes nelze oddělovat od genetiky a epigenetiky. Celogenomové asociační studie odhalily desítky genetických variant, které ovlivňují strukturu nebo funkci synapsí a zvyšují riziko vzniku psychiatrických onemocnění. Mnohé z těchto variant se nacházejí v genech kódujících synaptické proteiny, jako jsou neuroligin, neurexin nebo shank, jejichž mutace jsou spojeny s autismem a schizofrenií. Pochopení toho, jak tyto genetické změny ovlivňují synaptickou funkci na molekulární úrovni, je nezbytným předpokladem pro vývoj personalizované medicíny, která by mohla nabídnout léčbu šitou na míru konkrétnímu pacientovi.
Budoucnost léčby neurologických a psychiatrických onemocnění tedy leží z velké části v hlubším pochopení synaptické biologie. Kombinace pokročilých zobrazovacích metod, molekulárně biologických přístupů, genetiky a farmakologie vytváří předpoklady pro vývoj terapií, které budou cílit na konkrétní synaptické mechanismy s dosud nepředstavitelnou přesností. Synapse, tento mikroskopický prostor mezi dvěma neurony, se tak stává jedním z nejdůležitějších bojišť moderní medicíny.
Publikováno: 03. 05. 2026
Kategorie: Neurovědy


