Plant Physiology otevírá nové kolo náboru na pozice Assistant Features Editors pro rok 2027
31. 07. 2026
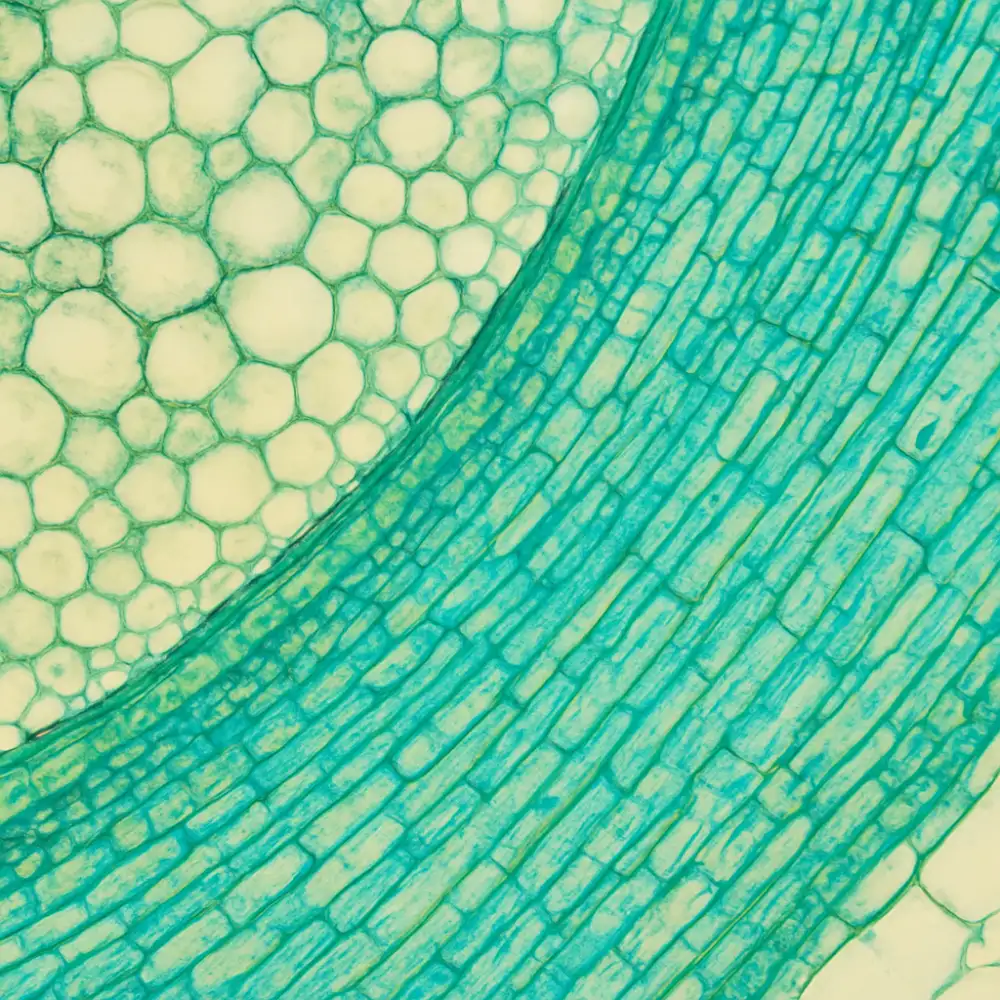
Rostlinná histologie představuje jednu z klíčových disciplín botaniky, která se zabývá studiem rostlinných tkání na mikroskopické úrovni. Jedná se o vědní obor, jenž zkoumá strukturu, složení, funkci a vzájemné vztahy buněk, které tvoří různé typy tkání v tělech rostlin. Histologie botanika, tedy rostlinná histologie, je neodmyslitelnou součástí moderní botaniky a poskytuje základní poznatky nezbytné pro pochopení fyziologie, morfologie a ekologie rostlin jako celku.
Samotný pojem histologie pochází z řeckého slova „histos, které znamená tkáň, a „logos, tedy věda nebo nauka. V kontextu botaniky se pak tato disciplína zaměřuje výhradně na rostlinné organismy a jejich tkáňové struktury, čímž se odlišuje od živočišné histologie, která studuje tkáně živočichů. Rostlinné tkáně jsou nesmírně rozmanité a jejich studium odhaluje fascinující svět mikroskopických struktur, jež jsou na první pohled neviditelné, přesto však zcela zásadní pro pochopení toho, jak rostliny fungují, rostou a přizpůsobují se podmínkám prostředí.
Význam rostlinné histologie nelze přeceňovat. Bez hlubokého pochopení tkáňové organizace rostlin by nebylo možné plně porozumět procesům, jako jsou fotosyntéza, transport vody a živin, mechanická podpora rostlinného těla nebo reprodukce. Každá tkáň v rostlině plní specifickou funkci a je tvořena buňkami s charakteristickými morfologickými a biochemickými vlastnostmi, které jsou přizpůsobeny právě té roli, jíž daná tkáň v organismu zastává.
Rostlinná histologie se dělí na několik základních oblastí podle typu studovaných tkání. Meristematické tkáně, tedy tkáně dělivé, jsou předmětem intenzivního zájmu, protože právě ony jsou zodpovědné za růst rostlin. Pletiva krycí, vodivá, základní a mechanická pak tvoří základ pro pochopení celkové architektury rostlinného těla. Studium těchto tkání umožňuje vědcům nejen porozumět normálnímu vývoji rostlin, ale také identifikovat patologické změny způsobené chorobami, škůdci nebo nepříznivými podmínkami prostředí.
V praktické rovině má rostlinná histologie obrovský přínos pro zemědělství, lesnictví, potravinářský průmysl i farmacii. Znalost tkáňové struktury rostlin umožňuje šlechtitelům vyvíjet odolnější a výnosnější odrůdy plodin, farmaceutům identifikovat léčivé látky a jejich lokalizaci v rostlinném těle a ekologům lépe chápat adaptační mechanismy rostlin na různé typy stanovišť. Histologické metody, jako je světelná mikroskopie, elektronová mikroskopie nebo histochemické barvení, patří mezi základní nástroje, bez nichž by moderní botanický výzkum nebyl myslitelný.
Rostlinná histologie tak stojí na pomezí několika vědních oborů a její poznatky prostupují celou botanickou vědou. Je to disciplína, která propojuje makroskopický pohled na rostlinu s mikroskopickým světem buněk a tkání, čímž vytváří ucelený obraz o fungování rostlinných organismů. Bez ní by naše chápání rostlinného života zůstalo neúplné a povrchní, neschopné postihnout hloubku a složitost biologických procesů, které se v rostlinách každý okamžik odehrávají.
Zájem o vnitřní stavbu rostlin sahá hluboko do minulosti, přestože skutečný vědecký výzkum rostlinných tkání byl podmíněn vývojem optických přístrojů, bez nichž by pozorování buněčných struktur nebylo možné. Již starověcí filozofové a lékaři se zajímali o stavbu rostlinných orgánů, avšak jejich poznání bylo omezeno na to, co bylo patrné pouhým okem. Teprve v sedmnáctém století nastal zlom, který položil základy moderní rostlinné histologie.
Robert Hooke, anglický přírodovědec a polyhistor, publikoval v roce 1665 své zásadní dílo Micrographia, v němž popsal pozorování tenkých řezů korku pod primitivním mikroskopem vlastní konstrukce. Hooke si všiml pravidelně uspořádaných dutých komůrek, které přirovnal k celám klášterů a nazval je latinsky „cellulae. Přestože Hooke pozoroval pouze odumřelé buněčné stěny bez živého obsahu, jeho objev je považován za jeden z nejvýznamnějších milníků v dějinách biologie vůbec. Tento moment byl přelomový i pro botanickou histologii, protože poprvé ukázal, že rostlinné orgány mají složitou vnitřní architekturu, která není viditelná pouhým okem.
Přibližně ve stejné době pracoval nizozemský obchodník a samouk Antonie van Leeuwenhoek, jehož mikroskopy dosahovaly pro tehdejší dobu nevídaného zvětšení. Leeuwenhoek popsal živé buňky v různých rostlinných i živočišných materiálech a jeho pozorování výrazně rozšířila chápání mikrosvěta. Paralelně s ním rozvíjel botanickou mikroskopii Nehemiah Grew, anglický lékař a botanik, který vydal v roce 1682 monumentální dílo The Anatomy of Plants. Grew v něm systematicky popsal vnitřní stavbu různých rostlinných orgánů, rozlišil různé typy pletiv a věnoval pozornost cévním svazkům, parenchymatickým buňkám i průduchům. Italský přírodovědec Marcello Malpighi pracoval ve stejném období a jeho dílo Anatome Plantarum z roku 1675 přineslo srovnatelně cenné poznatky. Malpighi a Grew jsou dnes považováni za zakladatele rostlinné anatomie jako vědecké disciplíny.
Osmnácté století přineslo spíše konsolidaci dosavadních poznatků než zásadní nové objevy, přestože zdokonalování mikroskopů pomalu pokračovalo. Skutečný průlom nastal až ve století devatenáctém, kdy Matthias Jakob Schleiden a Theodor Schwann formulovali buněčnou teorii. Schleiden v roce 1838 prohlásil, že všechny rostlinné organismy jsou složeny z buněk nebo jejich produktů, čímž dal botanické histologii pevný teoretický základ. Tato teorie, doplněná Schwannovými pozorováními živočišných tkání, změnila celou biologii a otevřela cestu systematickému studiu rostlinných pletiv.

Ve druhé polovině devatenáctého století se botanická histologie rychle rozvíjela díky zdokonalení mikroskopické techniky, zavedení fixačních metod a barvicích technik. Wilhelm Hofmeister přispěl zásadními poznatky o buněčném dělení a vývoji rostlinných embryí. Julius von Sachs, jeden z nejvýznamnějších botaniků devatenáctého století, systematizoval poznatky o rostlinných tkáních a zavedl terminologii, která se v modifikované podobě používá dodnes. Sachs rozlišil základní typy pletiv a popsal jejich funkce v kontextu celého rostlinného těla.
Na přelomu devatenáctého a dvacátého století přispěl k rozvoji botanické histologie Gottlieb Haberlandt, jehož dílo Physiologische Pflanzenanatomie z roku 1884 představovalo první systematický pokus o propojení struktury rostlinných pletiv s jejich fyziologickými funkcemi. Haberlandt jako první navrhl myšlenku, že izolované rostlinné buňky by mohly být pěstovány v umělém prostředí, čímž předjímal pozdější rozvoj technik tkáňových kultur. Tato vize se naplnila až ve dvacátém století, kdy se tkáňové kultury staly nepostradatelným nástrojem rostlinné biologie a šlechtitelství.
Dvacáté století přineslo do botanické histologie revoluční změny díky zavedení elektronové mikroskopie, která umožnila pozorovat ultrastrukturu buněk a odhalila detaily buněčných organel, buněčné stěny a plasmodezmat. Výzkumy Keithy Esau, americké botaničky původem z Ruska, vedly ke vzniku monumentálního díla Plant Anatomy, které se stalo standardní učebnicí rostlinné anatomie a histologie po celém světě. Esau popsala vývoj a strukturu cévních pletiv s dosud nevídanou přesností a její práce ovlivnila celé generace botaniků.
V současné době je botanická histologie neoddělitelně spojena s molekulární biologií, genetikou a vývojovou biologií. Moderní výzkum využívá konfokální mikroskopii, fluorescenční značení proteinů, in situ hybridizaci a celou řadu dalších sofistikovaných metod, které umožňují sledovat nejen statickou strukturu pletiv, ale i dynamické procesy jejich vývoje a diferenciace. Přesto základní principy, které formulovali průkopníci od Hookea přes Schleidena až po Esau, zůstávají v jádru oboru pevně zakotveny a tvoří nezbytný základ pro pochopení složité architektury rostlinného těla.
Rostlinná histologie, tedy věda zabývající se studiem tkání rostlin na mikroskopické úrovni, představuje fascinující oblast botaniky, která nám odhaluje neuvěřitelnou složitost a organizaci živých rostlinných organismů. Každá rostlina, ať už jde o drobnou bylinu nebo mohutný strom, je tvořena různými typy tkání, přičemž každá z nich plní specifické a nezastupitelné funkce, které společně zajišťují přežití a prosperitu celého organismu.
Mezi základní typy rostlinných tkání patří tkáně dělivé, krycí, vodivé, základní a mechanické. Každá z těchto skupin se vyznačuje charakteristickou stavbou buněk, jejich uspořádáním a samozřejmě i funkcí, kterou v rostlinném těle zastávají. Pochopení těchto tkání je klíčové nejen pro botaniky a biology, ale také pro zemědělce, zahradníky a všechny, kdo se zajímají o fungování rostlinné říše.
Dělivé tkáně, odborně nazývané meristémy, tvoří základ pro růst a vývoj rostliny. Jsou složeny z buněk, které si zachovávají schopnost neustálého dělení, čímž zajišťují produkci nových buněk pro ostatní tkáně. Rozlišujeme primární meristémy, které se nacházejí na vrcholcích stonků a kořenů a jsou zodpovědné za délkový růst rostliny, a sekundární meristémy, jako je kambium a felogen, jež umožňují tloustnutí stonku a kořene. Bez funkčních meristémů by rostlina nebyla schopna růstu ani obnovy poškozených částí. Meristematické buňky jsou charakteristické svou malou velikostí, tenkými buněčnými stěnami a velkými jádry, což jsou znaky typické pro intenzivně se dělící buňky.
Krycí tkáně tvoří ochranný obal celé rostliny a jsou prvním obranným valem proti nepříznivým vlivům vnějšího prostředí. Pokožka, neboli epidermis, pokrývá mladé části rostliny a je tvořena zpravidla jednou vrstvou buněk, jejichž vnější stěny jsou impregnovány kutinem, což vytváří nepropustnou kutikulu chránící rostlinu před nadměrným výparem vody. Na povrchu pokožky se nacházejí průduchy, specializované struktury tvořené dvěma svěracími buňkami, které regulují výměnu plynů a transpirace. Tato regulace je naprosto zásadní pro fotosyntézu i pro vodní hospodářství rostliny. U starších částí rostlin, zejména u dřevin, je pokožka nahrazena druhotnou krycí tkání zvanou periderm, která obsahuje korková pletiva tvořená odumřelými buňkami naplněnými suberinem.
Vodivé tkáně představují jeden z nejdůležitějších systémů v těle cévnatých rostlin. Xylém a floém jsou dvě hlavní složky vodivého systému, přičemž každá z nich transportuje jiné látky. Xylém, neboli dřevo, zajišťuje transport vody a v ní rozpuštěných minerálních látek z kořenů do nadzemních částí rostliny. Je tvořen odumřelými buňkami, tracheidami a cévami, jejichž buněčné stěny jsou zpevněny ligninem. Floém naproti tomu transportuje organické látky, především sacharózu vzniklou při fotosyntéze, z listů do ostatních částí rostliny. Floémové buňky jsou živé a jejich funkce závisí na metabolické aktivitě přilehlých průvodních buněk. Vodivé tkáně jsou uspořádány do cévních svazků, jejichž rozmístění v těle rostliny se liší podle toho, zda jde o jednoděložnou nebo dvouděložnou rostlinu.

Základní tkáně tvoří největší objem rostlinného těla a plní celou řadu různorodých funkcí. Parenchym je nejrozšířenějším typem základní tkáně a jeho buňky jsou živé, s tenkými buněčnými stěnami a schopností obnovit dělivou aktivitu v případě potřeby. Asimilační parenchym, nazývaný také chlorenchym, obsahuje velké množství chloroplastů a je hlavním místem fotosyntézy. Zásobní parenchym slouží k ukládání rezervních látek, jako jsou škrob, tuky nebo bílkoviny. Vodní parenchym, typický pro sukulentní rostliny, akumuluje vodu a pomáhá rostlině přežít v suchých podmínkách. Aerenchym, parenchym s velkými mezibuněčnými prostory, se vyskytuje u vodních a bahenních rostlin a usnadňuje výměnu plynů v prostředí s omezeným přístupem vzduchu.
Mechanické tkáně zajišťují pevnost a pružnost rostlinného těla. Kolenchym je tvořen živými buňkami se zesílenými rohovými nebo lamelárními buněčnými stěnami a poskytuje mechanickou oporu mladým, dosud rostoucím částem rostliny. Je typický pro řapíky listů a okraje stonků. Sklerenchym naproti tomu tvoří pevnější oporu a je složen z buněk s rovnoměrně zesílenými a lignifikovanými stěnami. Sklerenchymatická vlákna a sklereidy, neboli kamenné buňky, jsou odumřelé buňky, jejichž hlavní funkcí je mechanická ochrana a zpevnění. Sklereidy se nacházejí například v dužině hrušek, kde způsobují jejich charakteristickou zrnitou texturu, nebo v tvrdých skořápkách ořechů.
Pochopení struktury a funkce jednotlivých rostlinných tkání je základem pro studium fyziologie rostlin, jejich adaptací na různá prostředí i pro aplikovaný výzkum v zemědělství a biotechnologiích. Rostlinná histologie tak tvoří nepostradatelný základ celé moderní botaniky a bez jejích poznatků by nebylo možné plně porozumět životu a fungování rostlinné říše.
Každá buňka rostliny je jako písmeno v knize přírody – teprve když pochopíme strukturu pletiv, dokážeme číst celý příběh života ukrytý v stonku, listu či kořeni. Botanická histologie nás učí, že krása rostlin nespočívá jen v jejich vnější podobě, ale především v dokonalé organizaci jejich vnitřního světa.
Radoslav Pečinka
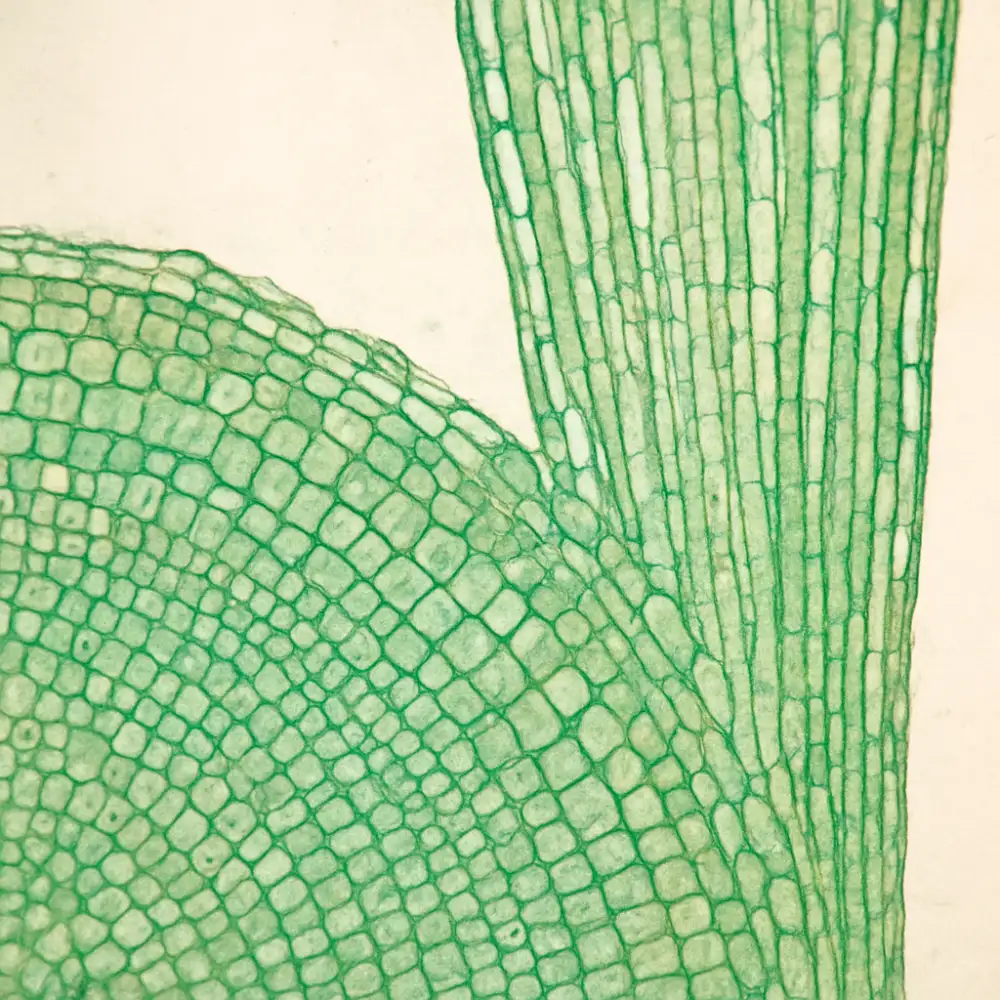
Rostlinná histologie, jakožto vědní disciplína zabývající se studiem rostlinných tkání na mikroskopické úrovni, věnuje zvláštní pozornost meristematickým tkáním, které představují základní stavební kameny veškerého rostlinného růstu. Meristémy jsou specializované oblasti rostlinného těla, v nichž dochází k intenzivnímu buněčnému dělení, a právě díky nim jsou rostliny schopny celoživotního růstu, což je výrazně odlišuje od většiny živočichů.
Buňky meristematických tkání mají velmi charakteristický vzhled, který je snadno rozpoznatelný při histologickém pozorování. Jsou to buňky s tenkými buněčnými stěnami, velkými jádry a hustou cytoplazmou, přičemž vakuoly jsou v nich přítomny jen v minimálním rozsahu nebo zcela chybí. Tato morfologie přímo odráží jejich fyziologickou funkci — neustálé dělení a produkci nových buněk, které se následně diferencují v různé typy specializovaných tkání.
Z hlediska botanické histologie rozlišujeme několik základních typů meristémů. Apikální meristémy se nacházejí na vrcholcích stonků a kořenů a jsou zodpovědné za takzvaný primární růst, tedy prodlužování rostlinného těla. Tyto struktury jsou ukryty v tzv. vegetačních vrcholech, kde jsou chráněny před mechanickým poškozením — u stonku listovými základy, u kořene kořenovou čepičkou. Histologické preparáty vegetačních vrcholů patří k nejkrásnějším a nejinformativnějším objektům rostlinné mikroskopie, protože na nich lze sledovat postupnou diferenciaci buněk od totipotentních iniciál až po plně specializované elementy cévních svazků, pokožky či základní tkáně.
Laterální meristémy, především kambium a felogen, zajišťují sekundární růst rostlin, tedy jejich tloustnutí. Kambium je tenká vrstva meristematických buněk uložená mezi dřevem a lýkem, a právě jeho aktivita vede ke každoroční tvorbě nových vrstev xylému a floému. Tento proces je dobře patrný na příčných řezech dřevnatými stonky, kde se projevuje jako letokruhy — jeden z nejznámějších příkladů toho, jak histologická struktura odráží biologické procesy probíhající v průběhu celého života stromu. Felogen neboli korkoví kambium pak produkuje ochranné korkové pletivo, které nahrazuje původní pokožku při druhotném tloustnutí orgánů.
Zvláštní kategorii tvoří interkalární meristémy, které jsou typické zejména pro jednoděložné rostliny, jako jsou trávy a obilniny. Tyto meristémy jsou uloženy v bazálních částech internodií a umožňují prodlužování stonku i po poranění jeho vrcholu. Právě díky nim dokáže tráva po posekání opět rychle dorůst — fenomén, který má obrovský ekologický i zemědělský význam.
Rostlinná histologie studuje nejen samotné meristematické buňky, ale také procesy, jimiž se z těchto buněk vyvíjejí různé typy trvalých tkání. Proces diferenciace buněk je řízen komplexní souhrou fytohormonů, zejména auxinů, cytokininů a giberellinů, přičemž každý z těchto hormonů ovlivňuje různé aspekty buněčného dělení, prodlužování a zrání. Auxiny produkované apikálními meristémy potlačují růst postranních pupenů — jev označovaný jako apikální dominance — a zároveň stimulují prodlužování buněk v subapikální zóně.
Histologické metody, jako je světelná mikroskopie fixovaných a barvených řezů, elektronová mikroskopie nebo imunohistochemické techniky, umožňují detailně sledovat strukturu a dynamiku meristematických tkání. Barvení hematoxylinem a eosinem, safraninem nebo toluidinovou modří odhaluje různé složky buněčných stěn a obsah buněk, čímž poskytuje cenné informace o stavu a aktivitě meristémů. Moderní metody zahrnují také in situ hybridizaci a konfokální mikroskopii, které umožňují lokalizovat expresi specifických genů přímo v tkáních vegetačního vrcholu.
Pochopení meristematických tkání má zásadní praktický význam pro zemědělství, zahradnictví i biotechnologie. Tkáňové kultury, při nichž se z malých explantátů obsahujících meristematické buňky regenerují celé rostliny, jsou dnes rutinně využívány k množení vzácných nebo obtížně množitelných druhů, k produkci zdravého sadebního materiálu prostého virových infekcí nebo k tvorbě geneticky modifikovaných rostlin. Meristematické buňky jsou pro tyto účely ideální právě proto, že si zachovávají totipotenci — schopnost vyvinout se v jakýkoli buněčný typ a za vhodných podmínek obnovit celý organismus.
Rostlinná histologie tak v oblasti meristematických tkání propojuje základní vědecký výzkum s praktickými aplikacemi a ukazuje, jak hluboké poznání mikroskopické struktury může přinést zcela konkrétní výsledky v podobě nových odrůd, efektivnějších pěstitelských postupů nebo lépe odolných plodin schopných čelit výzvám měnícího se klimatu.
Rostlinné tělo je neustále vystaveno celé řadě vnějších vlivů, které mohou narušit jeho integritu a ohrozit životně důležité procesy probíhající uvnitř pletiv. Právě proto příroda vyvinula během milionů let evoluce sofistikované systémy ochrany, které souhrnně označujeme jako krycí tkáně. Tyto tkáně tvoří hranici mezi vnitřním prostředím rostliny a okolním světem, přičemž jejich stavba i funkce jsou předmětem studia rostlinné histologie, tedy botanické histologie, která se zabývá mikroskopickou anatomií rostlinných orgánů a jejich pletiv.
Základní a nejrozšířenější krycí tkání je pokožka neboli epidermis. Tato jednovrstevná tkáň pokrývá povrch mladých rostlinných orgánů, jako jsou listy, stonky, kořeny a plody. Buňky epidermis jsou zpravidla těsně k sobě přiléhající, bez mezibuněčných prostor, což zajišťuje mechanickou pevnost a zároveň brání nežádoucímu úniku vody. Vnější stěny epidermálních buněk jsou zesíleny a na povrchu pokryty kutikulou, což je voskovitá vrstva tvořená kutinem. Tato vrstva hraje klíčovou roli v regulaci transpirace a chrání rostlinu před nadměrným výparem vody, před ultrafialovým zářením i před průnikem patogenních mikroorganismů.
Epidermis není monotónní vrstvou identických buněk. Naopak, v rámci tohoto pletiva se diferencují specializované buněčné typy, které plní různé funkce. Nejznámějšími jsou průduchy neboli stomata, tvořené dvěma svěracími buňkami, jež regulují výměnu plynů mezi rostlinou a atmosférou. Průduchy umožňují příjem oxidu uhličitého potřebného pro fotosyntézu a zároveň regulují výdej vodní páry při transpiraci. Jejich rozložení na povrchu listu, hustota i způsob otevírání a zavírání jsou přizpůsobeny životním podmínkám konkrétního druhu rostliny.
Dalším typem specializovaných buněk epidermis jsou trichomy, tedy výrůstky pokožky, které mohou mít podobu jednoduchých chloupků, žláznatých útvarů nebo šupin. Trichomy plní celou řadu ochranných funkcí. Mohou odrážet sluneční záření a tím snižovat přehřívání listů, mohou vylučovat silice nebo pryskyřice odpuzující herbivory, nebo naopak produkovat lepkavé látky zachycující hmyz, jako je tomu u masožravých rostlin. Hustý porost trichomů na povrchu listů může také vytvářet vzduchovou vrstvu, která zpomaluje transpiraci a chrání rostlinu před vysycháním.
S přibývajícím věkem rostliny a zejména u dřevin dochází k nahrazení primární epidermis sekundárním krycím pletivem, kterým je periderm. Periderm vzniká činností fellogenu, tedy korkového kambia, které produkuje směrem ven korková pletiva a směrem dovnitř feloderm. Korek neboli felém je tvořen odumřelými buňkami s buněčnými stěnami prosycenými suberinem, což je hydrofobní látka zabraňující průchodu vody a plynů. Díky tomu tvoří korek vynikající izolační vrstvu, která chrání vnitřní pletiva před mrazem, suchem, mechanickým poškozením i před napadením patogeny.

V korku se nacházejí čočinky neboli lenticely, které jsou funkčním ekvivalentem průduchů v epidermis. Prostřednictvím lenticel probíhá výměna plynů mezi vnitřními pletivy dřevin a atmosférou. Jejich přítomnost je dobře patrná na kůře mladých větví jako malé, často vyvýšené tečky nebo proužky. Struktura a rozmístění lenticel jsou druhově specifické a mohou sloužit jako diagnostický znak při určování dřevin.
Rostlinná histologie věnuje krycím tkáním mimořádnou pozornost, protože jejich stavba odráží adaptace rostlin na konkrétní prostředí. Xerofyty, tedy rostliny suchých stanovišť, mají silně vyvinutou kutikulu, hluboce zanořené průduchy a hustý porost trichomů. Hydrofyty naopak mají kutikulu redukovanou nebo zcela chybějící a průduchy jsou u plovoucích listů přesunuty na svrchní stranu čepele. Tyto morfologické a anatomické adaptace jsou výsledkem dlouhodobého evolučního tlaku a představují fascinující příklady toho, jak forma sleduje funkci.
Studium krycích tkání v kontextu botanické histologie tak přináší nejen poznatky o stavbě rostlinného těla, ale také hlubší pochopení vztahů mezi rostlinami a jejich prostředím, jejich obranných strategií a evolučních přizpůsobení, která umožnila rostlinám osídlit prakticky každý kout naší planety.
Rostlinná histologie se zabývá studiem tkání na mikroskopické úrovni a jednou z nejfascinujících oblastí tohoto vědního oboru je bezesporu problematika vodivých tkání. Tyto specializované struktury představují základ cévnatých rostlin a umožňují jim přežívat v nejrůznějších podmínkách prostředí. Xylém a floém jsou dvě základní vodivé tkáně, které společně tvoří cévní svazky a zajišťují transport látek po celém těle rostliny.
Xylém, někdy označovaný také jako dřevo, je tkáň primárně zodpovědná za transport vody a minerálních látek z kořenů směrem nahoru do listů a ostatních nadzemních částí rostliny. Tento transport probíhá na základě transpirační tenze, kapilárních sil a kořenového tlaku. Z histologického hlediska je xylém složen z několika typů buněk, přičemž nejdůležitější jsou tracheidy a cévní členy. Tracheidy jsou dlouhé, vřetenovité buňky s tečkovanými stěnami, které se vyskytují zejména u nahosemenných rostlin. Cévní členy jsou naproti tomu kratší a širší, s perforovanými příčnými stěnami, které umožňují přímý tok vody. V průběhu diferenciace buněk xylému dochází k odumírání protoplastů a ke ztluštění a lignifikaci buněčných stěn, čímž vznikají pevné, pevnostně odolné struktury schopné odolávat negativnímu tlaku vznikajícímu při transpiraci.
Vedle vodivých elementů obsahuje xylém také xylémová vlákna, která plní především mechanickou funkci, a xylémový parenchym, jehož buňky zůstávají živé a podílejí se na ukládání zásobních látek a na laterálním transportu vody a iontů. Primární xylém vzniká z prokambia a diferencuje se v průběhu primárního růstu rostliny, zatímco sekundární xylém, tvořící dřevo dřevin, vzniká činností kambia a přirůstá každoročně ve formě letokruhů. Studium letokruhů, tzv. dendrochronologie, umožňuje rekonstruovat klimatické podmínky v minulosti a datovat stáří dřevěných předmětů.
Floém je naproti tomu tkáň zajišťující transport organických látek, především asimilátů vzniklých fotosyntézou, z míst jejich produkce do míst spotřeby nebo ukládání. Tento transport je označován jako translokace a probíhá aktivním způsobem za spotřeby energie. Základními funkčními buňkami floému jsou sítkovicové buňky u nahosemenných rostlin a sítkovicové články u krytosemenných rostlin. Sítkovicové články jsou propojeny tzv. sítkovými deskami s četnými póry, jimiž proudí asimiláty. Na rozdíl od buněk xylému si sítkovicové články zachovávají živý protoplast, avšak postrádají jádro, což je kompenzováno těsnou funkční spoluprací s průvodními buňkami, které jsou metabolicky velmi aktivní a zásobují sítkovicové články potřebnou energií.
Průvodní buňky jsou charakteristickým prvkem floému krytosemenných rostlin a jejich přítomnost je jedním z histologických znaků, které odlišují tuto skupinu rostlin od ostatních. Průvodní buňky a sítkovicové články vznikají z téže mateřské buňky a jsou propojeny četnými plazmodezmatickými spojeními, která umožňují intenzivní výměnu látek a signálů. Floémový parenchym pak plní zásobní funkci a podílí se na nakládání a vykládání asimilátů v místech zdroje a spotřeby.
Z hlediska botanické histologie je velmi důležité rozlišovat primární a sekundární floém. Primární floém, stejně jako primární xylém, vzniká z prokambia a je přítomen u všech cévnatých rostlin. Sekundární floém vzniká činností kambia a je charakteristický pro dřeviny, přičemž jeho buňky mají kratší životnost než buňky sekundárního xylému a jsou postupně nahrazovány nově vznikajícím sekundárním floémem. Starší vrstvy sekundárního floému se mění v lýkovou část borky a plní ochrannou funkci.
Cévní svazky, v nichž jsou xylém a floém uspořádány společně, mohou mít různou stavbu v závislosti na systematickém zařazení rostliny a na části rostlinného těla, ve které se nacházejí. Kolaterální cévní svazky, v nichž floém leží na vnější straně xylému, jsou nejběžnějším typem a vyskytují se v listech a stoncích většiny dvouděložných rostlin. Bikolaterální svazky mají floém na obou stranách xylému a jsou typické například pro lilkovité rostliny. Radiální cévní svazky, charakteristické pro kořeny, mají xylém a floém uspořádány střídavě v radiálním směru.

Pochopení stavby a funkce vodivých tkání má zásadní význam nejen pro teoretickou botaniku, ale také pro praktické obory jako je zemědělství, zahradnictví a lesnictví. Znalost histologické stavby xylému a floému umožňuje lépe porozumět procesům příjmu vody a živin, translokaci asimilátů a mechanismům obrany rostlin proti patogenům, kteří velmi často napadají právě vodivé tkáně a způsobují závažná onemocnění vedoucí k vadnutí a odumírání rostlin.
Parenchymatické tkáně představují v rostlinné histologii naprosto zásadní skupinu tkání, která tvoří základ většiny rostlinných orgánů a zajišťuje celou řadu životně důležitých funkcí. Jedná se o tkáně složené z živých buněk, které si zpravidla zachovávají schopnost dělit se a přizpůsobovat se měnícím se podmínkám prostředí. Parenchym je považován za výchozí tkáň, z níž evolučně vznikly ostatní specializované tkáně rostlin. Tato skutečnost sama o sobě vypovídá o jeho výjimečném postavení v celém systému rostlinné histologie.
Z hlediska stavby jsou parenchymatické buňky typicky izodiametrické, tedy přibližně stejně velké ve všech směrech, ačkoliv tato charakteristika není absolutní a existují četné výjimky. Buněčná stěna parenchymatických buněk bývá tenká a primární, přičemž obsahuje četné plasmodesmy, které umožňují komunikaci mezi sousedními buňkami. Velká centrální vakuola zaujímá podstatnou část buněčného objemu a hraje klíčovou roli v udržování turgoru a v regulaci osmotického tlaku. Cytoplazma je přitlačena k buněčné stěně a jádro bývá relativně velké, což odráží metabolickou aktivitu těchto buněk.
Asimilační parenchym, označovaný také jako chlorenchym, představuje jednu z nejdůležitějších forem základního parenchymu. Právě v chlorenchymu probíhá fotosyntéza, tedy proces, na němž závisí prakticky veškerý život na Zemi. Buňky asimilačního parenchymu jsou bohaté na chloroplasty a nacházejí se především v listech, kde tvoří mezofyl. V listech dvouděložných rostlin rozlišujeme palisádový parenchym, jehož buňky jsou protáhlé a kolmo orientované k povrchu listu, a houbovitý parenchym, jehož buňky jsou nepravidelně uspořádané s výraznými mezibuněčnými prostorami. Toto uspořádání není náhodné – palisádový parenchym maximalizuje zachycení světelné energie, zatímco houbovitý parenchym usnadňuje výměnu plynů nezbytnou pro průběh fotosyntézy i dýchání.
Zásobní parenchym plní v rostlinném těle zcela odlišnou, avšak neméně důležitou funkci. Jeho buňky slouží k ukládání zásobních látek, mezi nimiž dominují škrob, tuky, bílkoviny a různé organické kyseliny. Zásobní parenchym se hojně vyskytuje v semenech, hlízách, oddenku, cibulích a dužnatých plodech, kde akumulované látky slouží jako energetická rezerva pro klíčení nebo vegetativní rozmnožování. Zvláštní formou zásobního parenchymu jsou buňky ukládající vodu, které se vyskytují u sukulentních rostlin přizpůsobených životu v aridních podmínkách. Tyto buňky mají obrovské vakuoly naplněné vodou a slizovitými látkami, které brání rychlému odpařování.
Vzdušný parenchym, nazývaný aerenchym, je specializovanou tkání, která se vyvinula jako adaptace na život ve vodním nebo zamokřeném prostředí. Aerenchym je charakterizován rozsáhlou sítí mezibuněčných prostor vyplněných vzduchem, které zajišťují přívod kyslíku do kořenů ponořených v anoxickém substrátu. Tato tkáň je typická pro rostliny mokřadů, jako jsou rákos, orobinec nebo lekníny. Mezibuněčné prostory aerenchymu mohou vznikat buď schizogenně, tedy rozestupováním buněk, nebo lysigenně, tedy rozpadem a odumíráním buněk.
Vodivý parenchym, přestože se svým názvem odlišuje od ostatních forem, je nedílnou součástí cévních svazků. Parenchymatické buňky obklopující floém a xylém se podílejí na nakládání a vykládání asimilátů a minerálních látek, čímž regulují transport látek v celém rostlinném těle. Buňky průvodní, které jsou funkčně spjaty se sítkovicemi floému, jsou typickým příkladem vysoce specializovaného parenchymu s intenzivním metabolismem.
Základní parenchym, který vyplňuje prostory mezi ostatními tkáněmi v různých orgánech rostlin, plní roli mechanické výplně a zároveň slouží jako místo pro ukládání různých metabolitů. V kořenech tvoří základní parenchym kortex, tedy kůru, která obklopuje centrální válec a podílí se na příjmu a transportu vody a minerálních látek z půdy. Endodermis, tvořená jednou vrstvou parenchymatických buněk s Casparyho proužky, představuje selektivní bariéru regulující průchod látek do cévního systému kořene.
Rostlinná histologie věnuje parenchymatickým tkáním mimořádnou pozornost také proto, že tyto tkáně jsou nositeli totipotence, tedy schopnosti diferencovat se v jakýkoliv jiný buněčný typ. Tato vlastnost je základem vegetativního rozmnožování, regenerace poraněných tkání i moderních biotechnologických postupů, jako je tkáňová kultura. Dedifikovaný parenchym může za vhodných podmínek vytvořit kalusovou tkáň, z níž lze vypěstovat celou novou rostlinu. Tato pozoruhodná plasticita parenchymatických buněk je jedním z nejvýznamnějších fenoménů celé rostlinné biologie a zůstává předmětem intenzivního výzkumu dodnes.
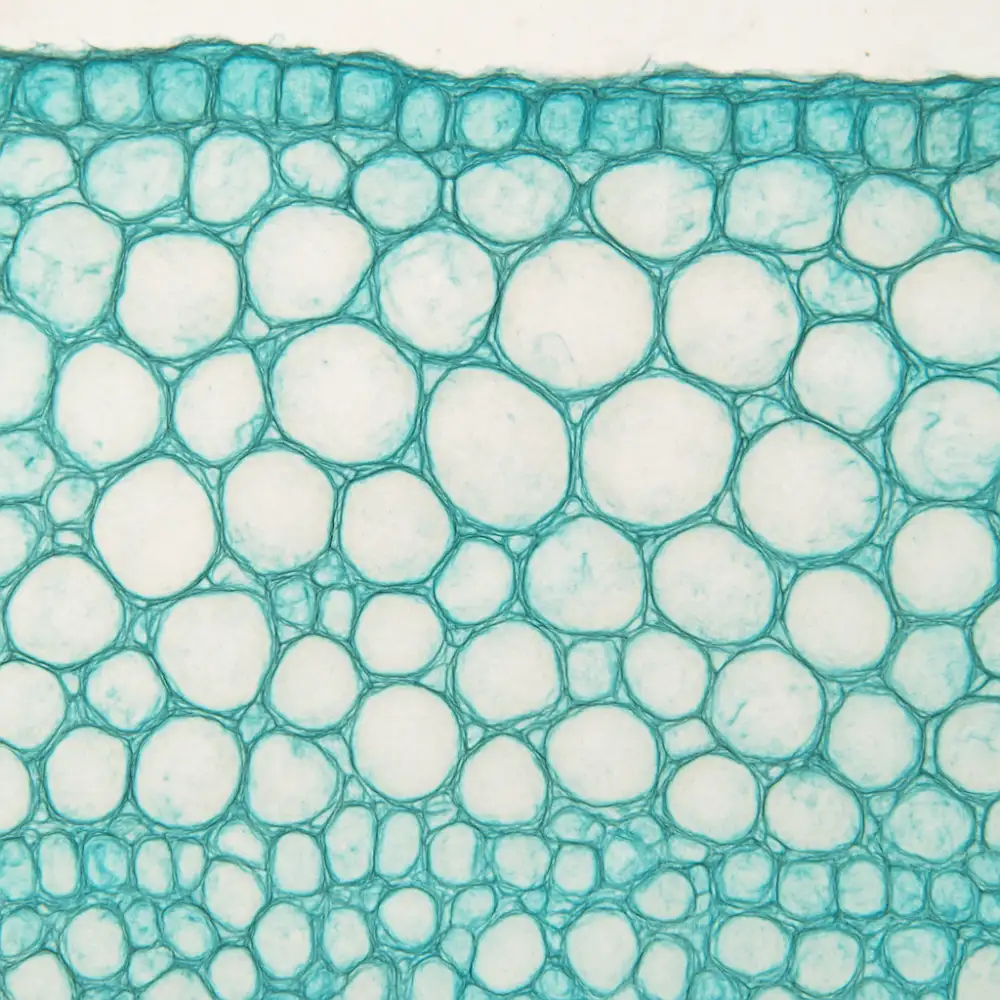
Rostlinná histologie, tedy věda zabývající se studiem rostlinných tkání na mikroskopické úrovni, věnuje značnou pozornost mechanickým tkáním, které jsou klíčové pro zajištění pevnosti a stability rostlinného těla. Bez těchto specializovaných tkání by rostliny nebyly schopny odolávat mechanickým silám, jako je tlak větru, tíha vlastního těla nebo působení gravitace. Mechanické tkáně tvoří jakýsi vnitřní skelet rostliny, který jí umožňuje udržet vzpřímený postoj a chránit citlivé vnitřní struktury před poškozením.
V botanické histologii rozlišujeme především dva základní typy mechanických tkání, a to kolenchym a sklerenchym. Každý z těchto typů má svou specifickou stavbu, funkci i umístění v těle rostliny, přičemž oba hrají nezastupitelnou roli v celkovém mechanickém systému rostlinného organismu.
Kolenchym je živá mechanická tkáň, která se vyskytuje především v mladých, ještě rostoucích částech rostliny. Jeho buňky jsou živé i v dospělém stavu a vyznačují se nerovnoměrně zesílenými buněčnými stěnami, které jsou tvořeny celulózou a pektinem. Tato zesílení nejsou rovnoměrně rozložena po celém obvodu buňky, ale soustřeďují se zpravidla v rozích nebo na tangenciálních stěnách. Právě díky tomu, že buněčné stěny kolenchymu neobsahují lignin, zůstává tato tkáň pružná a schopná dalšího růstu, což je zásadní vlastnost pro části rostliny, které ještě neukončily svůj vývoj. Kolenchym se nachází typicky těsně pod pokožkou stonků, v okrajích listů nebo v řapících, kde poskytuje pružnou oporu bez toho, aby bránil dalšímu prodlužování orgánů.
Sklerenchym naproti tomu představuje mechanickou tkáň tvořenou zpravidla odumřelými buňkami, jejichž buněčné stěny jsou silně zesíleny ligninem. Lignifikace buněčných stěn způsobuje, že tyto buňky ztrácejí schopnost dalšího růstu a jejich protoplast postupně odumírá, přičemž funkci přebírají právě pevné buněčné stěny. Sklerenchym se dělí na dva základní typy buněk, a to sklerenchymatická vlákna a skleridy. Sklerenchymatická vlákna jsou protáhlé buňky s velmi silnými lignifikovanými stěnami, které se sdružují do svazků a poskytují rostlině výjimečnou tahovou i tlakovou pevnost. Jsou hojně zastoupena ve stonkách dvouděložných rostlin, kde obklopují cévní svazky a tvoří pevné mechanické prstence nebo pásy. Skleridy jsou naproti tomu buňky různých tvarů, od izodiametrických kamenných buněk až po rozvětvené hvězdicovité formy. Kamenné buňky, odborně nazývané brachyskleridy, jsou typické například pro dužinu hrušek, kde způsobují jejich charakteristickou zrnitou konzistenci, nebo pro tvrdé skořápky peckovin.
Z hlediska rostlinné histologie je mimořádně zajímavé sledovat, jakým způsobem jsou mechanické tkáně rozmístěny v průřezu různými rostlinnými orgány. Rozložení mechanických tkání v rostlinném těle není náhodné, ale řídí se přesnými biomechanickými principy. Ve stonku jsou mechanické tkáně zpravidla soustředěny na periferii nebo tvoří rozptýlené svazky, čímž se maximalizuje odolnost vůči ohybu při minimálním množství materiálu. Tento princip je srovnatelný s inženýrskými řešeními, jako jsou duté trubky nebo nosníky s I-profilem, které také dosahují vysoké pevnosti při relativně nízké hmotnosti.
Rostlinná histologie dále zkoumá, jak se mechanické tkáně vyvíjejí v průběhu ontogeneze rostliny. Primární mechanické tkáně, tedy kolenchym a primární sklerenchym, vznikají z prokambia a základního meristému během primárního růstu rostliny. Sekundární mechanické tkáně, zejména sekundární dřevo bohaté na lignifikovaná vlákna, vznikají činností kambia při sekundárním tloustnutí. Právě sekundární dřevo, tedy xylém, představuje nejdůležitější mechanickou tkáň dřevin, která jim umožňuje dorůstat do obrovských rozměrů a odolávat extrémním mechanickým silám po celá staletí.
Studium mechanických tkání má také praktický význam pro pochopení vlastností rostlinných materiálů využívaných člověkem. Vlákna lnu, konopí, juty nebo bambusu jsou tvořena právě sklerenchymatickými vlákny, jejichž výjimečné mechanické vlastnosti jsou využívány v textilním průmyslu, stavebnictví i moderních kompozitních materiálech. Rostlinná histologie tak přesahuje rámec čistě akademické vědy a nachází uplatnění v celé řadě praktických oborů lidské činnosti.
Studium rostlinných buněk pomocí mikroskopu představuje jeden z nejzásadnějších pilířů botanické histologie, bez něhož by naše poznání vnitřní stavby rostlinných těl zůstalo velmi omezené. Rostlinná histologie jako vědní disciplína se opírá o celou řadu mikroskopických technik, které se v průběhu staletí vyvíjely a zdokonalovaly, přičemž každá z nich přináší specifický pohled na strukturu buněk, pletiv a orgánů rostlin.
Základním nástrojem, který stál u samotného zrodu botanické histologie, byl světelný mikroskop. Jeho princip spočívá v průchodu viditelného světla preparátem, přičemž čočkový systém zvětšuje obraz pozorovaného objektu. Světelná mikroskopie umožňuje sledovat buněčné stěny, vakuoly, chloroplasty a další organely, avšak její rozlišovací schopnost je fyzikálně omezena vlnovou délkou světla. Přesto zůstává světelný mikroskop v botanické histologii nezastupitelným nástrojem pro každodenní práci, neboť je relativně dostupný a umožňuje pozorovat živé buňky v jejich přirozeném stavu.
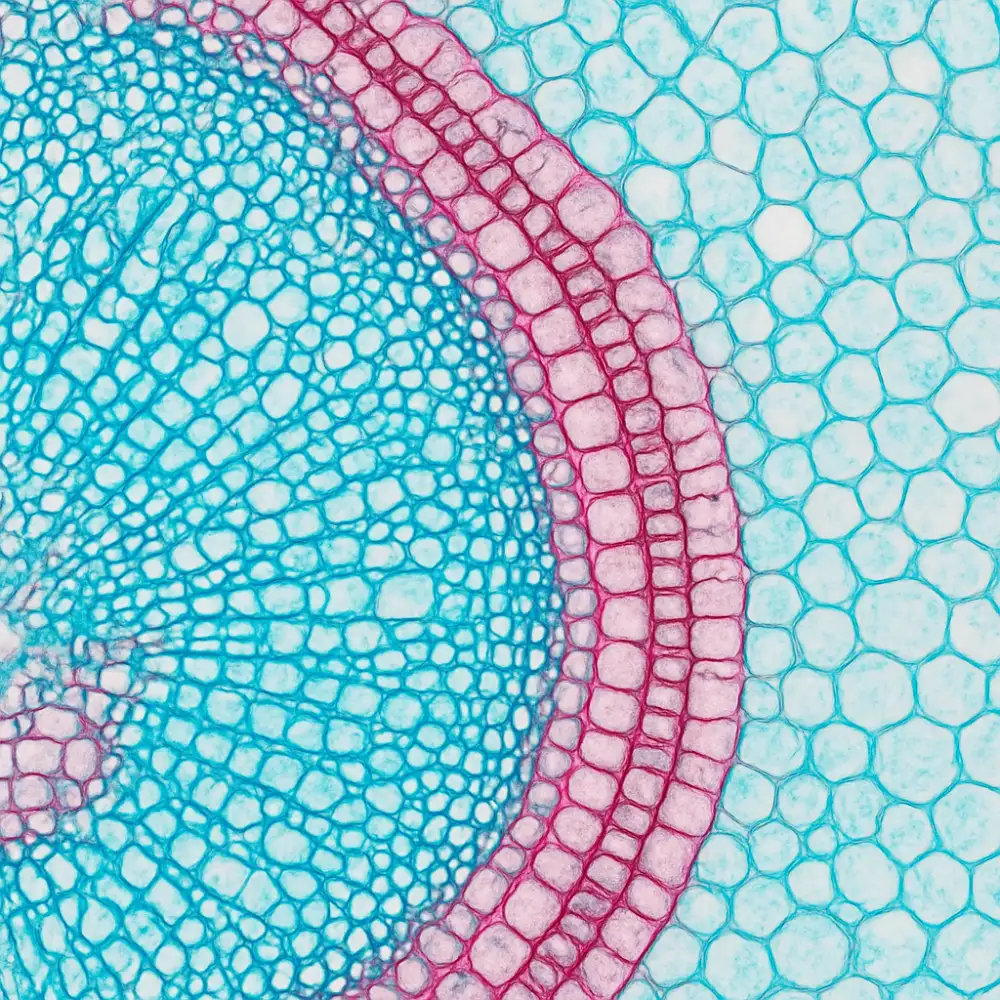
Pro přípravu kvalitních preparátů je nezbytné provést fixaci rostlinného materiálu, která zastaví veškeré biologické procesy a zachová struktury buněk v co nejpřirozenějším stavu. K fixaci se nejčastěji používají chemické látky jako formaldehyd, glutaraldehyd nebo směs FAA, tedy formaldehyd, kyselina octová a etanol. Po fixaci následuje odvodnění vzorku a jeho zalití do parafinu nebo pryskyřice, což umožňuje přípravu velmi tenkých řezů pomocí mikrotomu. Tloušťka řezů pro světelnou mikroskopii se obvykle pohybuje v rozmezí pěti až dvaceti mikrometrů, přičemž tenčí řezy poskytují lepší průhlednost a detailnější zobrazení buněčných struktur.
Samotné pozorování nezabarvených řezů by přineslo jen velmi omezené informace, proto se v botanické histologii hojně využívají různé histochemické barvicí metody. Safranin a Astrablau jsou klasickou kombinací, která odlišuje lignifikované buněčné stěny od celulózových. Safranin barví lignin a suberinizované stěny červeně, zatímco Astrablau obarvuje celulózu modrozeleně. Tato kombinace je nesmírně cenná při studiu cévních svazků, kde je nutné rozlišit xylém od floému. Lugolův roztok slouží k průkazu škrobu, který se po aplikaci tohoto reagens zbarví do tmavě modrofialova, což je zvláště užitečné při studiu zásobních pletiv. Fluorescenční barviva jako kalcofluor white umožňují vizualizaci celulózových vláken buněčné stěny při použití fluorescenčního mikroskopu, kde excitační světlo určité vlnové délky vyvolá emisi světla o jiné vlnové délce.
Konfokální laserová rastrovací mikroskopie představuje výrazný pokrok oproti klasické fluorescenční mikroskopii. Tato technika umožňuje získávat optické řezy vzorkem a rekonstruovat trojrozměrné obrazy buněčných struktur, aniž by bylo nutné fyzicky krájet preparát. V botanické histologii nachází konfokální mikroskopie uplatnění zejména při studiu buněčných stěn, plasmodesmat a dynamiky cytoskeletu. Plasmodesmata, tedy drobné cytoplazmatické kanálky propojující sousední buňky, jsou struktury, jejichž studium bylo ještě před několika desetiletími velmi obtížné, avšak moderní konfokální mikroskopie v kombinaci s fluorescenčními sondami umožňuje jejich vizualizaci s nebývalou přesností.
Elektronová mikroskopie posunula hranice poznání rostlinných buněk do zcela nové dimenze. Transmisní elektronový mikroskop pracuje s urychlovanými elektrony místo fotonů, čímž dosahuje rozlišení v řádu nanometrů, tedy tisíckrát lepšího než světelný mikroskop. Díky transmisní elektronové mikroskopii bylo možné poprvé detailně popsat ultrastrukturu chloroplastů, mitochondrií, endoplazmatického retikula a Golgiho aparátu v rostlinných buňkách. Příprava vzorků pro transmisní elektronovou mikroskopii je však velmi náročná a zahrnuje fixaci glutaraldehydem a osmium tetroxidem, odvodnění, zalití do epoxidové pryskyřice a přípravu ultratenkých řezů o tloušťce přibližně šedesáti až devadesáti nanometrů. Kontrastování řezů uranyl acetátem a citrátem olova zvyšuje kontrast zobrazených struktur, neboť elektrony jsou různými materiály různě rozptylovány.
Rastrovací elektronová mikroskopie naopak umožňuje studovat povrchy rostlinných orgánů a buněk s mimořádnou plasticitou a hloubkou ostrosti. Tato technika nachází uplatnění při studiu povrchu listů, struktury průduchů, trichomů a povrchu pylových zrn. Morfologie povrchu rostlinných orgánů, studovaná rastrovací elektronovou mikroskopií, přináší cenné taxonomické informace a pomáhá při determinaci rostlinných druhů.
Atomová silová mikroskopie je relativně mladou technikou, která umožňuje zobrazovat povrchy biologických vzorků s atomárním rozlišením bez nutnosti jejich vakuování nebo kovového napařování. V botanické histologii se tato metoda využívá zejména při studiu architektury buněčné stěny a uspořádání celulózových mikrofibril. Pochopení mechanických vlastností buněčné stěny je klíčové pro porozumění růstu rostlinných buněk a diferenciaci pletiv.
Rostlinná histologie se zabývá studiem tkání na mikroskopické úrovni a jedním z nejzásadnějších témat, které tato disciplína zkoumá, je bezesporu buněčná stěna jako strukturní prvek zcela odlišující rostlinné buňky od buněk živočišných. Zatímco živočišné buňky jsou obklopeny pouze tenkou plazmatickou membránou, rostlinné buňky mají navíc pevnou ochrannou vrstvu, která jim propůjčuje mechanickou stabilitu, tvar a celou řadu dalších vlastností nezbytných pro přežití a fungování celého organismu.
Buněčná stěna není jen pasivní obal. Jde o dynamickou strukturu, která se v průběhu života buňky mění, přizpůsobuje a specializuje podle toho, jakou funkci daná buňka v rámci tkáně plní. Rostlinná histologie rozlišuje několik typů buněčných stěn, přičemž základní dělení vychází z toho, zda se jedná o stěnu primární nebo sekundární. Primární buněčná stěna se tvoří jako první, v době, kdy buňka ještě roste a dělí se. Je poměrně tenká, pružná a umožňuje buňce zvětšovat svůj objem. Skládá se především z celulózy, hemicelulózy a pektinů, přičemž tyto složky jsou uspořádány do složité sítě, která je schopna odolávat mechanickému namáhání, aniž by ztrácela svou elasticitu.

Jakmile buňka dosáhne svého konečného tvaru a přestane růst, může začít ukládat sekundární buněčnou stěnu, která se tvoří mezi primární stěnou a plazmatickou membránou. Tato vrstva je výrazně silnější a tužší, neboť obsahuje větší množství celulózy a navíc bývá prostoupena ligninem, látkou, která celé struktuře dodává mimořádnou pevnost. Lignifikace buněčné stěny je klíčovým procesem při vzniku dřevnatých tkání, jako jsou xylémová vlákna nebo tracheidy, a bez tohoto procesu by stromy nebyly schopny dorůstat do výšek, které běžně pozorujeme v přírodě.
Z pohledu botanické histologie je fascinující sledovat, jak se buněčná stěna liší v různých typech rostlinných tkání. V parenchymatických buňkách, které tvoří základ většiny základních tkání, je stěna tenká a primární, zatímco v kolenchymu je nerovnoměrně ztlustlá, zejména v rozích buněk, což tomuto pletivovému typu propůjčuje schopnost mechanické opory při zachování určité pružnosti. Sklerenchym naproti tomu disponuje buňkami s velmi silnou, zpravidla lignifikovanou sekundární stěnou, přičemž samotná buňka v dospělosti většinou odumírá a zanechává za sebou pouze pevnou schránku plnící funkci výztuže.
Buněčná stěna rovněž hraje nezastupitelnou roli v transportních procesech uvnitř rostlinného těla. Skrze drobné kanálky zvané plazmodesmy procházejí spojení mezi sousedními buňkami, což umožňuje přenos látek a signálů napříč tkáněmi. Tyto mikroskopické struktury jsou součástí buněčné stěny a jejich přítomnost nebo absence výrazně ovlivňuje, jak intenzivně spolu buňky komunikují a jak efektivně probíhá transport vody, živin a asimilátů.
Nelze opomenout ani ochrannou funkci buněčné stěny vůči vnějším patogenům. Rostliny nemají imunitní systém ve smyslu, jak ho chápeme u živočichů, a proto je buněčná stěna jejich první a velmi důležitou obrannou linií. Při napadení houbou nebo bakterií dochází k rychlému zpevnění stěny a ukládání specifických látek, které brání proniknutí patogenu do nitra buňky. Rostlinná histologie tyto obranné reakce studuje na tkáňové úrovni a pomáhá odhalit, jakými mechanismy se rostliny brání a jak lze tyto mechanismy využít v zemědělství nebo šlechtitelství.
Celkově vzato, buněčná stěna představuje jeden z nejdůležitějších a nejcharakterističtějších znaků rostlinné buňky, a její studium v rámci botanické histologie přináší zásadní poznatky nejen o stavbě a funkci jednotlivých tkání, ale také o evoluci rostlin, jejich adaptacích na různá prostředí a o procesech, které umožňují rostlinám dosáhnout tak obdivuhodné rozmanitosti forem a strategií přežití.
Rostlinná histologie prošla v posledních desetiletích naprosto zásadní proměnou, a to především díky nástupu moderních zobrazovacích a analytických technologií, které vědcům umožňují nahlížet do struktury rostlinných pletiv způsoby, jež byly ještě před několika generacemi nepředstavitelné. Klasické metody, jako bylo barvení řezů hematoxylinem nebo safraninem a jejich pozorování pod světelným mikroskopem, sice stále tvoří základ výuky i základního výzkumu, ale moderní věda je dnes daleko za tímto horizontem.
| Typ pletiva | Latinský název | Funkce | Tloušťka buněčné stěny | Přítomnost chloroplastů | Typická velikost buněk (µm) | Místo výskytu v rostlině | Živé buňky v dospělosti |
|---|---|---|---|---|---|---|---|
| Epidermis | Epidermis | Ochrana povrchu | Tenká až středně silná | Zpravidla ne (kromě průduchů) | 20–50 | Povrch listů, stonků, kořenů | Ano |
| Palisádový parenchym | Parenchyma palissadicum | Fotosyntéza | Tenká | Ano (40–50 chloroplastů/buňka) | 30–80 | Svrchní vrstva listu | Ano |
| Houbový parenchym | Parenchyma spongiosum | Výměna plynů, fotosyntéza | Tenká | Ano (méně než palisádový) | 20–60 | Spodní vrstva listu | Ano |
| Kolenchym | Collenchyma | Mechanická opora mladých orgánů | Nerovnoměrně ztlustlá | Výjimečně | 50–200 | Okraje listů, mladé stonky | Ano |
| Sklerenchym | Sclerenchyma | Pevnost a opora | Velmi silná (lignifikovaná) | Ne | 10–500 | Stonky, semena, plody | Ne (odumřelé) |
| Xylém | Xylem | Vedení vody a minerálních látek | Silná (lignifikovaná) | Ne | 20–500 | Cévní svazky celé rostliny | Ne (odumřelé cévy) |
| Floém | Phloem | Vedení organických látek (cukrů) | Tenká | Ne | 15–100 | Cévní svazky celé rostliny | Ano (sítkovice) |
| Meristém | Meristema | Dělení a růst rostliny | Velmi tenká | Proplastidy (nezralé) | 5–20 | Vrcholy kořenů a stonků, kambium | Ano |
Konfokální laserová rastrovací mikroskopie představuje jeden z největších průlomů v oblasti rostlinné histologie. Tato technika umožňuje vytváření trojrozměrných obrazů buněčných struktur s mimořádnou přesností a rozlišením. Díky ní mohou botanici sledovat rozmístění specifických proteinů, buněčných organel nebo dokonce průběh mitózy přímo v živých buňkách, aniž by bylo nutné tkáň fixovat nebo jinak poškozovat. Fluorescenční značení pomocí zelených fluorescenčních proteinů, takzvaných GFP, otevřelo zcela nové možnosti vizualizace dynamických procesů v rostlinných pásmech, jako je například transport auxinu nebo šíření signálů v cévních svazcích.

Dalším revolučním nástrojem je elektronová mikroskopie, a to jak transmisní, tak rastrovací. Transmisní elektronová mikroskopie dovoluje pozorovat ultrastrukturu buněčných stěn, plasmodesmat nebo chloroplastů s rozlišením v řádu nanometrů. Tímto způsobem bylo možné popsat detailní stavbu buněčné stěny, odhalit strukturu lamel v chloroplastech nebo vizualizovat endoplazmatické retikulum v jeho přirozené podobě. Rastrovací elektronová mikroskopie zase poskytuje nádherné trojrozměrné snímky povrchů listů, epidermálních buněk nebo pylových zrn, přičemž každý takový snímek odhaluje morfologické detaily, které jsou pro klasifikaci i funkční pochopení rostlinných struktur naprosto neocenitelné.
Imunohistochemické metody přinesly do rostlinné histologie možnost lokalizovat specifické molekuly přímo v tkáních. Pomocí protilátek konjugovaných s fluorescenčními barvivy nebo enzymy lze přesně určit, kde se v pletivech nacházejí konkrétní proteiny, polysacharidy nebo lipidy. Tato technika je obzvláště cenná při studiu buněčných stěn, kde umožňuje rozlišit různé typy pektinů, hemicelulóz nebo celulózy a sledovat jejich distribuci v závislosti na vývojovém stadiu buňky.
V posledních letech se do botanické histologie výrazně prosadily také metody molekulární biologie, zejména in situ hybridizace. Tato metoda umožňuje vizualizovat expresi konkrétních genů přímo v histologických řezech, a tak propojit morfologické pozorování s molekulárními daty. Vědci mohou například sledovat, ve kterých buňkách se exprimují geny zodpovědné za syntézu lignínu, nebo kde dochází k aktivaci genů řídících diferenciaci cévních pletiv.
Nezanedbatelný přínos přinesla také průtoková cytometrie, která sice není histologickou metodou v pravém slova smyslu, ale v kombinaci s histologickými přístupy umožňuje kvantifikovat obsah DNA v buňkách, analyzovat buněčný cyklus nebo třídit buněčné populace podle specifických povrchových markerů. Tato technika nachází uplatnění například při studiu polyploidie nebo při analýze buněčné diferenciace v meristémech.
Moderní zobrazovací technologie nezůstaly pozadu ani v oblasti automatizace a digitalizace. Digitální patologie a celoslajdové skenování umožňují vytvářet vysoce rozlišené digitální obrazy celých histologických preparátů, které lze následně analyzovat pomocí sofistikovaných softwarových nástrojů. Algoritmy strojového učení a umělé inteligence jsou dnes schopny automaticky identifikovat a klasifikovat různé typy buněk a pletiv, měřit jejich rozměry nebo detekovat patologické změny. Tím se výrazně zvyšuje objektivita a reprodukovatelnost histologické analýzy, která byla v minulosti do značné míry závislá na subjektivním úsudku pozorovatele.
Atomová silová mikroskopie přinesla možnost studovat mechanické vlastnosti buněčných stěn na nanometrické úrovni. Tato technika umožňuje měřit tuhost, elasticitu nebo adhezivní vlastnosti povrchů buněk a buněčných stěn, což je klíčové pro pochopení mechanismů růstu buněk a morfogeneze rostlinných orgánů. Propojení mechanických dat s histologickými pozorováními otevírá nové perspektivy v pochopení toho, jak fyzikální síly ovlivňují vývoj rostlinných pletiv.
Nelze opomenout ani pokroky v oblasti kryoelektronové tomografie, která umožňuje zobrazovat biologické vzorky v jejich přirozeném hydratovaném stavu bez nutnosti chemické fixace nebo kontrastování. Tato metoda přináší obrazy buněčných struktur v podmínkách blízkých živé buňce, čímž eliminuje artefakty vznikající při klasickém zpracování vzorků. Pro rostlinnou histologii má tento přístup mimořádný potenciál, protože buněčné stěny a vakuoly jsou struktury, které jsou při klasickém zpracování vzorků náchylné k deformacím.

Celkově lze říci, že moderní technologie zcela zásadně proměnily tvář rostlinné histologie. Integrace zobrazovacích, molekulárních a výpočetních metod umožňuje dnes vědcům studovat rostlinná pletiva s nebývalou hloubkou a přesností, přičemž každá nová technologie přináší nové otázky i nové odpovědi o tom, jak jsou rostliny stavěny a jak fungují.
Rostlinná histologie, tedy vědecká disciplína zabývající se studiem rostlinných tkání na mikroskopické úrovni, představuje jeden z nejdůležitějších nástrojů moderního zemědělství. Bez hlubokého porozumění tomu, jak jsou rostlinné tkáně organizovány, jak fungují a jak reagují na různé vnější i vnitřní podněty, by bylo prakticky nemožné dosahovat vysokých výnosů a zároveň udržovat zdraví pěstovaných plodin.
Jedním z klíčových praktických uplatnění rostlinné histologie je šlechtění rostlin. Šlechtitelé využívají poznatky o stavbě pletiv k tomu, aby mohli cíleně vybírat jedince s žádoucími vlastnostmi. Například tloušťka kutikuly, tedy voskové vrstvy pokrývající epidermální buňky, přímo ovlivňuje odolnost rostliny vůči suchu a různým patogenům. Pokud šlechtitel ví, jak tato tkáň vypadá u odolných odrůd, může pomocí histologických metod rychle identifikovat slibné kandidáty pro další křížení, aniž by musel čekat na výsledky polních pokusů trvajících několik let.
V oblasti ochrany rostlin hraje histologie botanika naprosto nezastupitelnou roli. Když zemědělec nebo agronomický poradce pozoruje na listech nebo stoncích podezřelé změny, histologická analýza umožňuje přesně určit, v jaké vrstvě tkáně se problém nachází a jaký je jeho původ. Rozlišení mezi houbovým onemocněním, bakteriální infekcí nebo poškozením způsobeným hmyzem je na základě histologického preparátu podstatně přesnější než pouhé vizuální hodnocení. Tato přesnost pak umožňuje zvolit správný postup ochrany a vyhnout se zbytečnému plošnému používání pesticidů, což je v dnešní době nesmírně důležité jak z ekonomického, tak z ekologického hlediska.
Rostlinná histologie nachází uplatnění také při studiu příjmu živin a vody. Cévní svazky, tvořené xylémem a floémem, jsou hlavními dálnicemi pro transport látek v rostlinném těle, a jejich histologická analýza odhaluje, zda jsou tyto dráhy funkční nebo zda jsou poškozeny například v důsledku nedostatku určitého minerálního prvku. Zemědělci tak mohou na základě histologických nálezů přizpůsobit hnojení a závlahu konkrétním potřebám plodiny v daném vývojovém stadiu.
Velký praktický význam má histologie rostlin také při hodnocení kvality osiva. Struktura semenného obalu, endospermu a zárodku je přímým ukazatelem životaschopnosti semene a jeho schopnosti vyklíčit za různých podmínek. Histologické metody umožňují odhalit skryté poškození, které není patrné pouhým okem, ale které by v praxi vedlo k výraznému snížení klíčivosti a tím i k ekonomickým ztrátám.
Dalším oblastí, kde se poznatky rostlinné histologie přímo promítají do zemědělské praxe, je pěstování zeleniny a ovoce pro potravinářský průmysl. Struktura parenchymatických buněk v plodech ovlivňuje jejich texturu, šťavnatost a trvanlivost. Výrobci potravin a pěstitelé spolupracují s histology na tom, aby vybrali odrůdy, jejichž tkáňová stavba zajistí co nejdelší skladovatelnost bez ztráty kvality. To má přímý dopad na snížení potravinových ztrát, které jsou celosvětově obrovským problémem.
Rostlinná histologie přispívá také k rozvoji precizního zemědělství. Moderní technologie umožňují automatizovanou analýzu histologických preparátů pomocí strojového učení, takže zemědělci mohou získávat detailní informace o stavu tkání svých plodin rychleji než kdykoli dříve. Tato propojení mezi klasickou histologií a moderními digitálními technologiemi otevírají zcela nové možnosti pro optimalizaci zemědělské produkce.
Nelze opomenout ani využití histologických poznatků při práci s tkáňovými kulturami. In vitro množení rostlin, které je dnes běžnou součástí produkce sadebního materiálu u mnoha plodin, by bez znalosti histologie nebylo možné. Pochopení toho, jak se diferencují meristematická pletiva a jak probíhá organogeneze, umožňuje optimalizovat podmínky kultivace a dosahovat vysoké úspěšnosti při regeneraci celých rostlin z malých explantátů.
Celkově lze říci, že bez rostlinné histologie by moderní zemědělství nemohlo fungovat na té úrovni efektivity a udržitelnosti, která je dnes vyžadována. Každý pokrok v porozumění mikroskopické stavbě rostlinných tkání se dříve či později promítne do konkrétních opatření na poli, ve skleníku nebo v laboratoři šlechtitelské stanice.
Publikováno: 21. 01. 2026
Kategorie: Botanika


