Pseudotěhotné myši v biomedicíně: nedostatek samic v estru zdražuje výzkum
12. 07. 2026
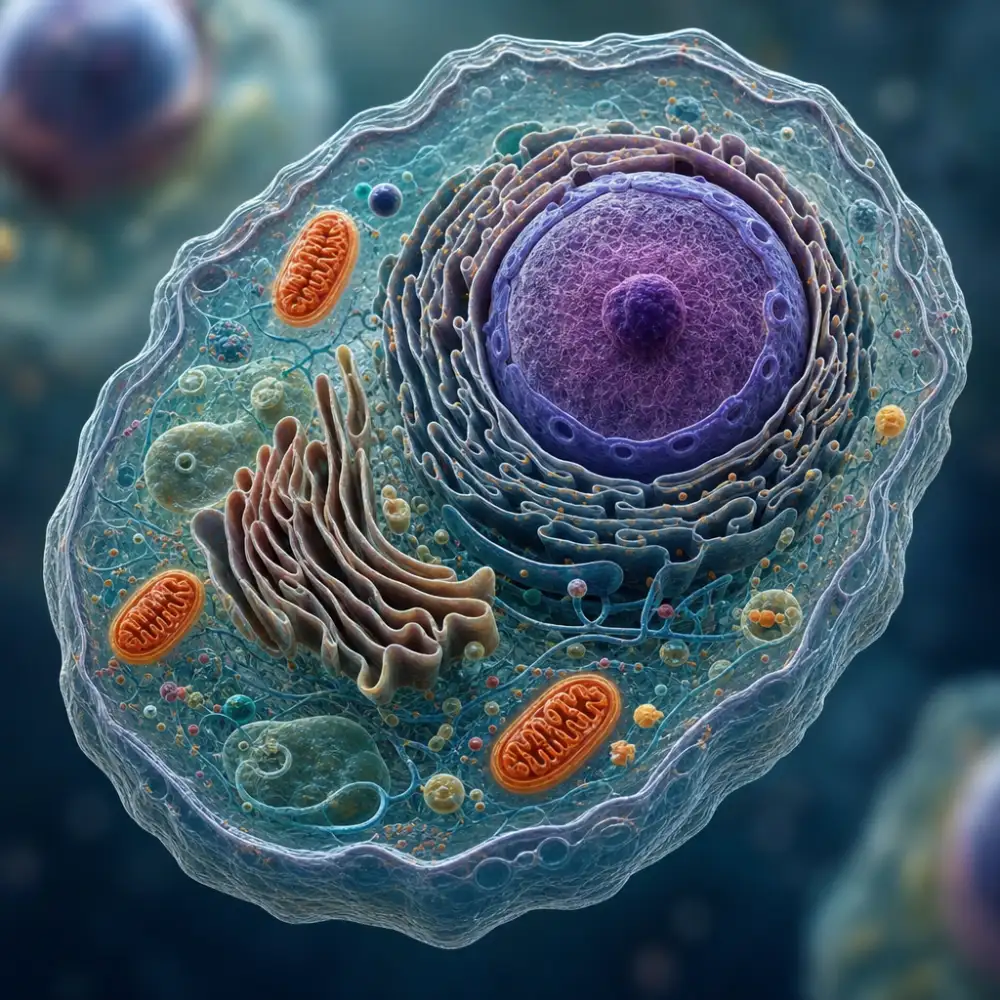
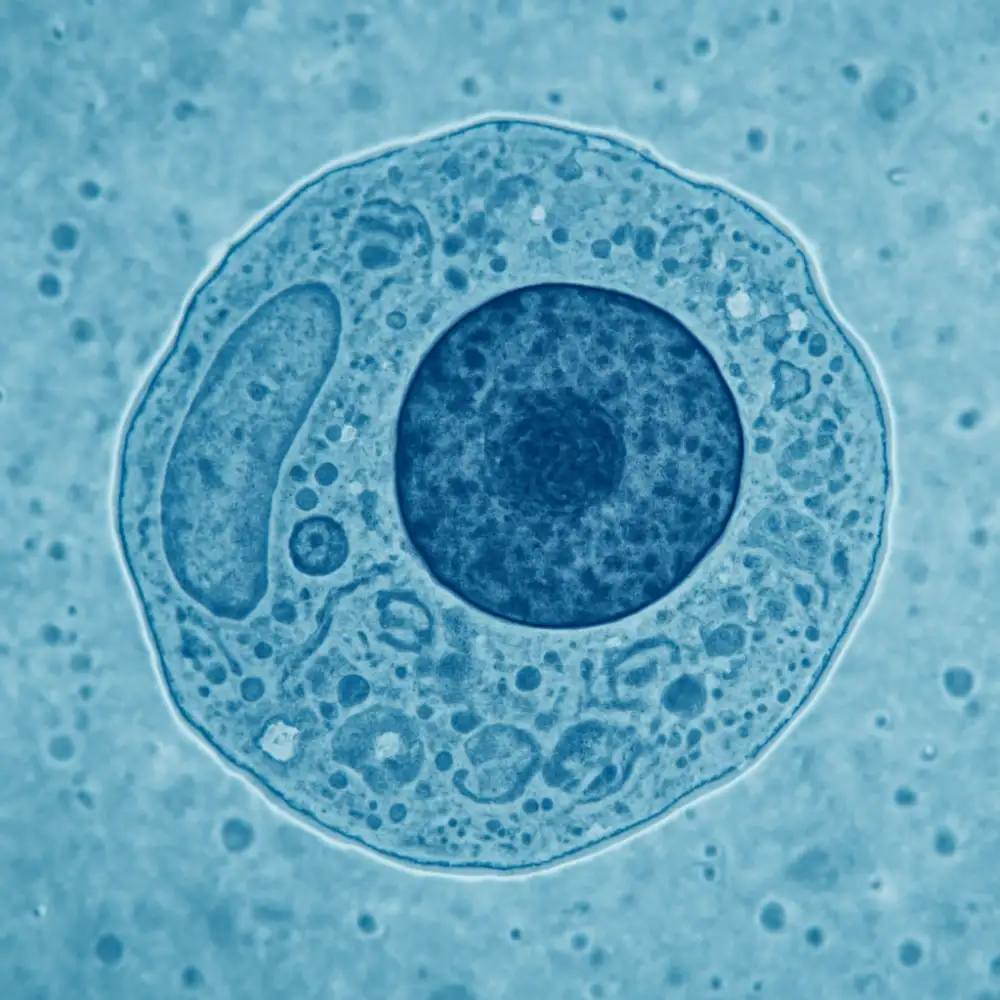
Eukaryotická buňka představuje základní stavební a funkční jednotku všech vyšších organismů, tedy rostlin, živočichů, hub a prvoků. Na rozdíl od jednodušších prokaryotických buněk se vyznačuje přítomností membránových organel a pravého buněčného jádra, ve kterém je uložena genetická informace ve formě DNA. Tato komplexní organizace umožňuje eukaryotickým buňkám vykonávat mnohem složitější funkce a dosahovat větších rozměrů než jejich prokaryotické protějšky.
Základní charakteristikou eukaryotické buňky je její kompartmentalizace, což znamená rozdělení vnitřního prostoru na jednotlivé oddíly ohraničené membránami. Toto uspořádání umožňuje současný průběh různých biochemických procesů v odlišných podmínkách, aniž by se navzájem rušily. Buněčná membrána tvoří vnější hranici buňky a reguluje transport látek mezi vnitřním a vnějším prostředím. Skládá se z fosfolipidové dvojvrstvy s vloženými proteiny, které plní nejrůznější funkce od transportu až po buněčnou komunikaci.
Buněčné jádro je nepochybně nejdůležitější organelou eukaryotické buňky. Je ohraničeno dvojitou jadernou membránou s jadernými póry, které kontrolují výměnu molekul mezi jádrem a cytoplazmatickým prostorem. Uvnitř jádra se nachází chromatin, který je tvořen DNA navinutou kolem histonových proteinů. V období buněčného dělení se chromatin kondenzuje do viditelných chromozomů. Jádro také obsahuje jadérko, strukturu specializovanou na syntézu ribozomální RNA a sestavování podjednotek ribozomů.
Cytoplazma představuje veškerý obsah buňky mimo jádro a je vyplněna cytosol, gelovou tekutinou obsahující rozpuštěné látky, enzymy a organely. V cytoplazmě se nacházejí ribozomy, které nejsou ohraničeny membránou a slouží jako místa syntézy proteinů. Eukaryotické ribozomy jsou větší než prokaryotické a mohou se vyskytovat volně v cytoplazmě nebo být připojeny k endoplazmatickému retikulu.
Endoplazmatické retikulum tvoří rozsáhlou síť membránových kanálků a cisteren prostupujících celou buňkou. Rozlišujeme drsné endoplazmatické retikulum, které má na povrchu připojeny ribozomy a podílí se na syntéze a modifikaci proteinů, a hladké endoplazmatické retikulum bez ribozomů, které se účastní syntézy lipidů a detoxikace látek. Golgického aparát navazuje na činnost endoplazmatického retikula a zajišťuje další úpravy, třídění a transport proteinů a lipidů k jejich cílovým destinacím.
Mitochondrie jsou organely s dvojitou membránou, které zajišťují buněčné dýchání a produkci energie ve formě ATP. Vnitřní membrána mitochondrií vytváří četné záhyby zvané kristy, které zvětšují povrch pro enzymatické reakce. Zajímavostí je, že mitochondrie obsahují vlastní DNA a ribozomy, což podporuje teorii o jejich původu z endosymbiózy s bakteriemi.
Rostlinné buňky navíc obsahují chloroplasty, organely zodpovědné za fotosyntézu, při které se světelná energie přeměňuje na chemickou energii uloženou v organických látkách. Chloroplasty mají také dvojitou membránu a obsahují vlastní genetický materiál. Lysozomy a peroxizomy jsou membránové váčky obsahující trávicí enzymy, které se podílejí na rozkladu makromolekul a detoxikaci škodlivých látek.
Cytoskelet tvoří vnitřní podpornou strukturu buňky složenou z proteinových vláken různých typů. Mikrotubuly, mikrofilamenta a intermediární filamenta zajišťují mechanickou podporu, umožňují pohyb organel a podílejí se na buněčném dělení. Tento dynamický systém umožňuje buňce měnit tvar a reagovat na vnější podněty.
Buněčné jádro představuje nejvýznamnější organelu eukaryotických buněk, která se vyznačuje přítomností dvojité membrány a obsahuje naprostou většinu genetické informace organismu. Tato struktura je charakteristická pro všechny eukaryotické organismy, tedy rostliny, živočichy, houby i prvoky, a odlišuje je od prokaryotických organismů, jako jsou bakterie a archeony, které jádro postrádají.
Genetická informace uložená v buněčném jádře je zakódována v molekulách deoxyribonukleové kyseliny, známé pod zkratkou DNA. Tato makromolekula má podobu dlouhých vláken, která jsou v jádře organizována do kompaktních struktur nazývaných chromozomy. Každý chromozom obsahuje jednu dlouhou molekulu DNA, která je úzce spojena s různými proteiny, zejména histony, jež pomáhají DNA sbalit do prostorově úsporné formy. Počet chromozomů je pro každý druh specifický, například člověk má v každé somatické buňce čtyřicet šest chromozomů uspořádaných do dvaceti tří párů.
Jaderná membrána, která obklopuje buněčné jádro, není jednoduchá bariéra, ale složitý systém dvou membrán s mezibrannným prostorem. Vnější membrána jaderného obalu je často spojena s endoplazmatickým retikulem a může být pokryta ribozomy. Jaderné póry jsou specializované proteinové komplexy, které prostupují oběma membránami a umožňují řízenou výměnu látek mezi jádrem a cytoplazmatickým prostorem buňky. Tyto póry jsou nezbytné pro transport molekul RNA z jádra do cytoplazmatu, kde probíhá syntéza proteinů, a zároveň umožňují vstup proteinů a dalších molekul do jádra.
Uvnitř jaderného prostoru se nachází nukleoplazma, gelová substance obsahující rozpuštěné ionty, enzymy a další molekuly nezbytné pro funkci jádra. V této oblasti probíhají klíčové procesy spojené s genetickou informací, především replikace DNA, transkripce a počáteční fáze zpracování RNA. Replikace DNA je proces, při kterém se vytváří identické kopie celého genomu, což je nezbytné před každým buněčným dělením, aby mohly obě dceřiné buňky získat kompletní genetickou výbavu.
Transkripce představuje přepis genetické informace z DNA do molekul RNA. Tento proces je řízen enzymy nazývanými RNA polymerázy, které čtou sekvenci DNA a syntetizují komplementární řetězec RNA. Vznikající molekuly messenger RNA neboli mRNA nesou informaci o pořadí aminokyselin v proteinech a po svém zpracování putují jaderními póry do cytoplazmatu, kde slouží jako předloha pro syntézu proteinů na ribozomech.
Jadérko je výrazná struktura viditelná uvnitř buněčného jádra, která není ohraničena membránou. Tato oblast je místem intenzivní syntézy ribozomální RNA a počátečního sestavování ribozomálních podjednotek. Jadérko se formuje kolem specifických úseků DNA obsahujících geny pro ribozomální RNA a jeho velikost odráží metabolickou aktivitu buňky.
Organizace genetické informace v jádře není náhodná. Chromozomy zaujímají v jaderném prostoru specifické oblasti nazývané chromozomová teritoria, přičemž aktivně transkribované oblasti DNA jsou často umístěny blíže k jaderným pórům. Tato prostorová organizace má zásadní význam pro regulaci genové exprese a koordinaci buněčných procesů.
Endoplazmatické retikulum představuje rozsáhlý membránový systém, který prostupuje celou eukaryotickou buňkou a tvoří složitou síť kanálků, cisteren a váčků. Tato organela je přímo napojena na jadernou membránu a hraje klíčovou roli v mnoha buněčných procesech, zejména pak v syntéze proteinů. Struktura endoplazmatického retikula se dělí na dva základní typy, které se liší jak svým vzhledem pod elektronovým mikroskopem, tak především svými funkcemi.
Drsné endoplazmatické retikulum získalo svůj název díky přítomnosti ribozomů na povrchu jeho membrán. Tyto ribozomy jsou zodpovědné za syntézu proteinů, které jsou určeny buď pro sekreci z buňky, nebo pro začlenění do membrán různých organel. Proces syntézy proteinů na drsném endoplazmatickém retikulu začíná v cytoplazmě, kde volné ribozomy zahájí translaci mRNA. Jakmile vznikající polypeptidový řetězec obsahuje signální sekvenci, specifickou posloupnost aminokyselin, je rozpoznán signálním rozpoznávacím proteinem.
Tento protein zastaví dočasně syntézu a nasměruje celý komplex ribozomu s rostoucím polypeptidem k membráně endoplazmatického retikula. Zde se ribosom připojí k specifickému receptoru a signální sekvence proteinu se provlékne přes proteinový kanál zvaný translokon přímo do lumenu endoplazmatického retikula. Během tohoto procesu pokračuje translace a nově vznikající protein se postupně dostává do vnitřního prostoru organely.
Uvnitř endoplazmatického retikula probíhají důležité posttranslační modifikace, které jsou nezbytné pro správné fungování proteinů. Jednou z nejdůležitějších modifikací je glykosylace, tedy připojování sacharidových řetězců k určitým aminokyselinám v proteinu. Tyto sacharidové skupiny jsou důležité pro správné skládání proteinu, jeho stabilitu a následné rozpoznávání v buňce. Enzymy přítomné v lumenu endoplazmatického retikula katalyzují také tvorbu disulfidických můstků mezi cysteinovými zbytky, což je kritické pro stabilizaci terciární struktury mnoha proteinů.
V endoplazmatickém retikulu funguje také sofistikovaný systém kontroly kvality, který zajišťuje, že pouze správně složené proteiny budou transportovány dále do Golgiho aparátu. Nesprávně složené nebo poškozené proteiny jsou rozpoznány speciálními chaperonovými proteiny a následně jsou buď znovu složeny, nebo označeny pro degradaci. Tento proces je známý jako endoplazmatická retikulum asociovaná degradace a chrání buňku před akumulací nefunkčních proteinů.
Hladké endoplazmatické retikulum neobsahuje na svém povrchu ribozomy a jeho funkce se liší od drsné varianty. Podílí se především na syntéze lipidů, metabolismu sacharidů a detoxikaci různých látek. V buňkách jater je hladké endoplazmatické retikulum obzvláště rozvinuté, protože zde probíhá intenzivní detoxikace škodlivých látek pomocí enzymu cytochromu P450.
Transport proteinů z endoplazmatického retikula do Golgiho aparátu probíhá prostřednictvím transportních vezikul, které se oddělují od membrány endoplazmatického retikula a následně se spojují s membránou Golgiho aparátu. Tento proces je přísně regulován a zajišťuje, že proteiny se dostanou na správné místo v buňce. Celý systém endoplazmatického retikula tak představuje základní pilíř pro správnou funkci eukaryotické buňky a její schopnost produkovat a distribuovat proteiny nezbytné pro život.
Golgiho aparát představuje klíčovou membránovou organelu eukaryotických buněk, která hraje nezastupitelnou roli v procesu transportu a modifikace různých látek uvnitř buňky. Tato složitá struktura byla poprvé popsána italským lékařem a vědcem Camillem Golgim na konci devatenáctého století, po němž následně získala své jméno. Golgiho aparát se nachází v cytoplazmě buňky, obvykle v blízkosti jádra a endoplazmatického retikula, se kterým úzce spolupracuje při zpracování proteinů a lipidů.
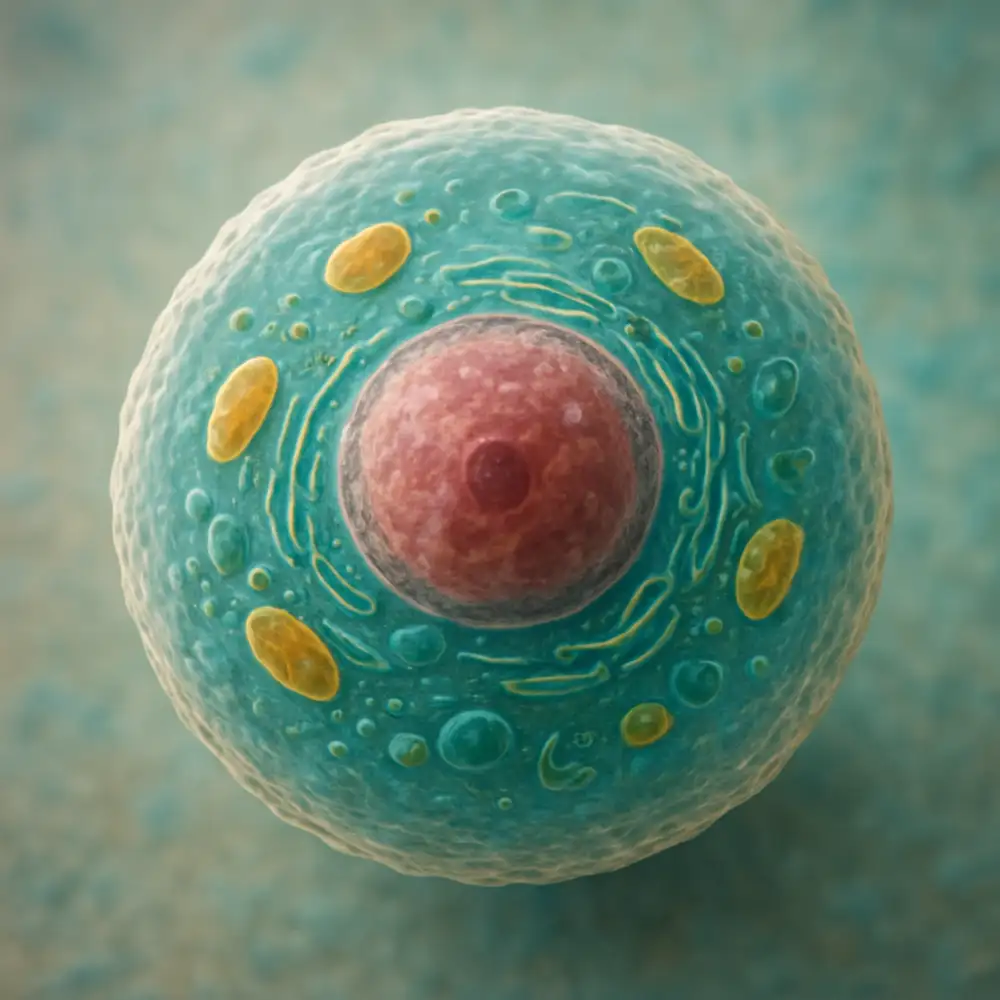
Struktura Golgiho aparátu je charakteristická svým uspořádáním do plochých membránových váčků nazývaných cisterny, které jsou uspořádány nad sebou podobně jako stohy talířů. Tyto cisterny nejsou izolované, ale tvoří funkční celek, který je polarizovaný. Rozlišujeme cis stranu, která je obrácená k endoplazmatickému retikulu a přijímá z něj transportní vezikuly, a trans stranu, ze které odcházejí vezikuly s hotovými produkty směrem k jejich konečnému cíli v buňce nebo mimo ni. Mezi těmito dvěma stranami se nachází střední oblast nazývaná mediální Golgi, kde probíhá většina modifikačních procesů.
Transport látek přes Golgiho aparát začína příjmem vezikul z drsného endoplazmatického retikula na cis straně. Tyto vezikuly obsahují nově syntetizované proteiny, které ještě nejsou plně funkční a vyžadují další úpravy. Proces postupného přesunu látek přes jednotlivé cisterny Golgiho aparátu umožňuje postupnou modifikaci molekul, která zahrnuje především glykosylaci, tedy přidávání sacharidových řetězců k proteinům a lipidům. Tato modifikace je kritická pro správnou funkci mnoha proteinů a jejich rozpoznávání jinými molekulami v buňce.
Během průchodu Golgiho aparátem dochází k sérii enzymatických reakcí, které postupně mění strukturu glykoproteinů. V cis části Golgiho aparátu se odstraňují některé cukry přidané v endoplazmatickém retikulu a začínají se připojovat nové. Ve střední části probíhá další modifikace sacharidových řetězců, zatímco v trans části se dokončuje glykosylace a proteiny jsou tříděny podle jejich konečného určení. Tento proces je nesmírně přesný a každý protein musí projít správnou sekvencí modifikací, aby získal svou finální funkční formu.
Golgiho aparát také hraje klíčovou roli v tvorbě lysozomů, které jsou důležité pro intracelulární trávení. Enzymy určené pro lysozomy jsou v Golgiho aparátu označeny specifickým značením, nejčastěji manóza-6-fosfátem, který slouží jako adresa zajišťující jejich správné doručení. Kromě lysozomů Golgiho aparát také produkuje sekreční vezikuly obsahující látky určené k vyloučení z buňky, jako jsou hormony, neurotransmitery nebo trávicí enzymy.
Mechanismus transportu mezi jednotlivými cisternami Golgiho aparátu je předmětem intenzivního vědeckého výzkumu. Existují dvě hlavní teorie vysvětlující tento transport. První teorie, nazývaná model vezikul, předpokládá, že cisterny zůstávají statické a látky se mezi nimi přenášejí pomocí malých transportních vezikul. Druhá teorie, model maturace cisteren, navrhuje, že samotné cisterny se postupně přeměňují z cis na trans formu a pohybují se přes Golgiho aparát, zatímco enzymy jsou recyklovány zpět pomocí vezikul. Současné důkazy naznačují, že pravděpodobně fungují oba mechanismy současně v závislosti na typu transportovaného nákladu.
Transport vezikul z Golgiho aparátu k jejich cílovým destinacím vyžaduje složitý systém molekulárního rozpoznávání a řízení. Proteiny na povrchu vezikul, nazývané SNARE proteiny, interagují s komplementárními proteiny na cílové membráně, čímž zajišťují specifické fúze vezikul pouze s příslušnými strukturami. Tento mechanismus zabraňuje chaosu v buňce a zajišťuje, že každá látka dorazí přesně tam, kde je potřeba.
Mitochondrie představují jedny z nejdůležitějších organel v eukaryotických buňkách a jejich role jako energetického centra je naprosto klíčová pro život všech vyšších organismů. Tyto drobné struktury, často připomínající tvar fazole nebo tyčinky, jsou zodpovědné za produkci hlavního energetického nosiče buňky, kterým je adenosintrifosfát neboli ATP. Bez fungujících mitochondrií by eukaryotické buňky nebyly schopny zajistit dostatečné množství energie pro své metabolické procesy a celý organismus by nemohl přežít.
Struktura mitochondrií je mimořádně složitá a dokonale přizpůsobená jejich funkci. Každá mitochondrie je obklopena dvěma membránami, vnější a vnitřní, přičemž vnitřní membrána vytváří četné záhyby nazývané kristy. Tyto kristy významně zvětšují povrch vnitřní membrány, což umožňuje umístění většího množství enzymatických komplexů podílejících se na výrobě energie. Prostor mezi oběma membránami se nazývá mezimembránový prostor, zatímco vnitřní část mitochondrie je označována jako matrix. V matrix se nacházejí enzymy Krebsova cyklu, mitochondriální DNA a ribozomy, což naznačuje částečnou autonomii těchto organel.
Proces tvorby energie v mitochondriích probíhá prostřednictvím buněčného dýchání, které se skládá z několika navazujících etap. První fází je glykolýza, která však probíhá v cytoplazmě buňky mimo mitochondrie. Při glykolýze dochází k rozštěpení molekuly glukózy na dvě molekuly pyruvátu za současného zisku malého množství ATP. Pyruvát následně vstupuje do mitochondrií, kde je přeměněn na acetyl-koenzym A, který vstupuje do Krebsova cyklu. Krebsův cyklus, známý také jako citrátový cyklus, představuje sérii enzymatických reakcí, při nichž dochází k postupné oxidaci acetyl-CoA a uvolňování elektronů, které jsou zachycovány nosiči NADH a FADH2.
Nejdůležitější fází produkce energie je však oxidativní fosforylace, která probíhá na vnitřní mitochondriální membráně. Během tohoto procesu elektrony z NADH a FADH2 procházejí řetězcem proteinových komplexů, známým jako dýchací řetězec nebo elektronový transportní řetězec. Při průchodu elektronů dýchacím řetězcem dochází k pumpování protonů z matrix do mezimembránového prostoru, čímž se vytváří elektrochemický gradient. Tento gradient představuje formu potenciální energie, která je následně využita enzymem ATP syntázou k fosforylaci ADP na ATP. Tento mechanismus, nazývaný chemiosmotická teorie, byl objeven Peterem Mitchellem a představuje jeden z nejvýznamnějších objevů v oblasti buněčné biologie.
Mitochondrie však nejsou pouze pasivními výrobci energie, ale aktivně reagují na potřeby buňky. Jejich počet v jednotlivých buňkách se může výrazně lišit v závislosti na energetických nárocích konkrétního typu buňky. Například svalové buňky nebo neurony obsahují tisíce mitochondrií, protože jejich metabolické požadavky jsou mimořádně vysoké. Naopak buňky s nižšími energetickými nároky obsahují mitochondrií méně. Mitochondrie jsou také schopny měnit svůj tvar, pohybovat se v buňce a dokonce se dělit nebo splývat, což umožňuje optimální distribuci energie v různých částech buňky.
Zajímavým aspektem mitochondrií je jejich evoluční původ. Podle endosymbiotické teorie vznikly mitochondrie z volně žijících bakterií, které byly pohlceny předchůdci eukaryotických buněk. Tato teorie je podporována několika důkazy, včetně přítomnosti vlastní kruhové DNA v mitochondriích, podobnosti mitochondriálních ribozomů s bakteriálními ribozomy a schopnosti mitochondrií dělit se binárním dělením podobně jako bakterie. Mitochondriální DNA kóduje pouze malou část proteinů potřebných pro funkci mitochondrie, zatímco většina genů byla během evoluce přenesena do jaderného genomu.
Poruchy funkce mitochondrií mohou vést k vážným onemocněním, známým jako mitochondriální choroby. Tyto poruchy postihují především tkáně s vysokými energetickými nároky, jako jsou svaly, nervový systém a srdce. Mitochondrie také hrají důležitou roli v procesu stárnutí buněk a v programované buněčné smrti neboli apoptóze, což podtrhuje jejich význam daleko přesahující pouhou produkci energie.
Lysozomy představují klíčové organely eukaryotické buňky, které jsou nepostradatelné pro rozklad a recyklaci buněčného materiálu. Tyto membránové váčky obsahují více než padesát různých typů hydrolytických enzymů, které jsou schopny štěpit prakticky všechny typy biologických makromolekul. Kyselé prostředí uvnitř lysozomu, udržované na hodnotě pH přibližně 4,5 až 5,0, vytváří optimální podmínky pro činnost těchto trávicích enzymů a zároveň chrání cytoplazmu před jejich destruktivním působením v případě narušení lysozomální membrány.
Proces trávení buněčného materiálu v lysozomech probíhá prostřednictvím několika mechanismů. Heterofagie představuje proces, při kterém buňka přijímá a rozkládá materiál z vnějšího prostředí. Tento mechanismus začíná endocytózou, kdy buňka pohlcuje extracelulární částice nebo tekutiny. Vzniklé endozomy následně splývají s lysozomy, čímž dochází ke vzniku sekundárních lysozomů, kde probíhá vlastní enzymatické štěpení přijatého materiálu. Produkty rozkladu, jako jsou aminokyseliny, monosacharidy nebo nukleotidy, jsou poté transportovány přes lysozomální membránu zpět do cytoplazmy, kde mohou být využity pro syntézu nových molekul nebo jako zdroj energie.
Autofagie představuje další významný mechanismus lysozomálního trávení, při kterém buňka degraduje vlastní poškozené nebo nepotřebné struktury. Tento proces je obzvláště důležitý při buněčném stárnutí, hladovění nebo při odstranění nefunkčních organel. Během autofagie dochází k obklopení cytoplazmatických komponent dvojitou membránou, což vede ke vzniku autofagozomu. Tento váček následně splývá s lysozomem a uzavřený materiál je enzymaticky rozložen. Autofagie tak umožňuje buňce přežít v nepříznivých podmínkách tím, že recykluje vlastní komponenty a získává z nich živiny.
Lysozomální enzymy jsou syntetizovány v drsném endoplazmatickém retikulu a následně modifikovány v Golgiho aparátu, kde získávají specifický marker v podobě manóza-6-fosfátu. Tento marker zajišťuje správné směrování enzymů do lysozomů. Transportní vezikuly obsahující nově syntetizované enzymy se oddělují od trans-Golgiho sítě a splývají s endozomy nebo přímo s lysozomy, čímž doplňují jejich enzymatickou výbavu.
Funkce lysozomů je nezbytná pro udržení buněčné homeostázy. Při poruchách lysozomálních enzymů dochází k hromadění nerozložených substrátů v buňce, což vede k závažným metabolickým onemocněním známým jako lysozomální choroby. Tyto dědičné poruchy demonstrují kritický význam lysozomálního systému pro normální fungování eukaryotických buněk. Lysozomy také hrají důležitou roli v imunitní odpovědi, kdy fagocyty využívají lysozomální enzymy k destrukci pohlcených bakterií a dalších patogenů, čímž přispívají k ochraně organismu před infekcemi.
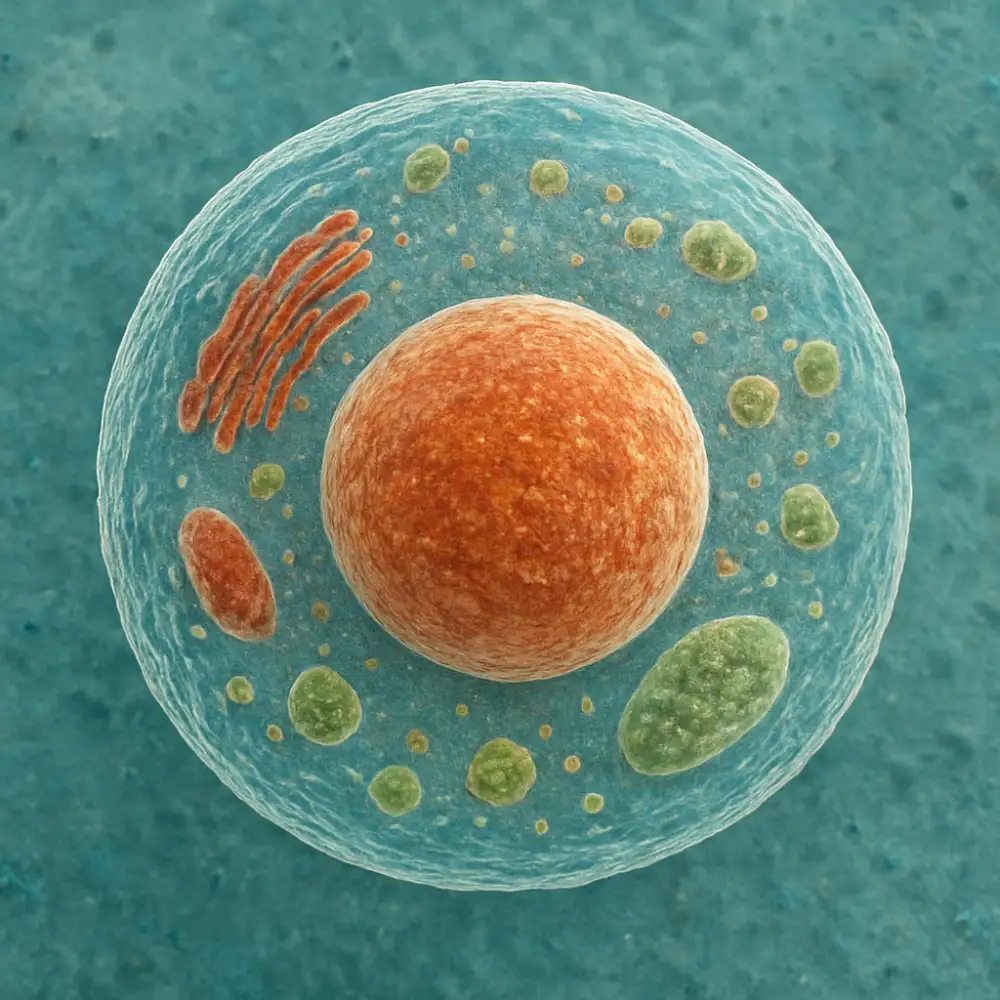
Cytoskelet představuje komplexní trojrozměrnou síť proteinových vláken, která prostupuje celou eukaryotickou buňkou a zajišťuje její strukturální integritu, tvar a umožňuje různé formy buněčného pohybu. Tento dynamický systém není statickou strukturou, ale neustále se přestavuje a reorganizuje v závislosti na aktuálních potřebách buňky. Cytoskelet se skládá ze tří hlavních typů proteinových filament, které se liší svou tloušťkou, chemickým složením i funkcí.
| Vlastnost | Eukaryotická buňka | Prokaryotická buňka |
|---|---|---|
| Jádro | Přítomno, ohraničené jadernou membránou | Chybí, DNA volně v cytoplazmě |
| Velikost buňky | 10-100 μm | 1-10 μm |
| DNA organizace | Lineární chromosomy s histony | Kruhová DNA bez histonů |
| Mitochondrie | Přítomny | Chybí |
| Endoplazmatické retikulum | Přítomno | Chybí |
| Golgiho aparát | Přítomen | Chybí |
| Ribozomy | 80S (větší) | 70S (menší) |
| Buněčné dělení | Mitóza a meióza | Binární dělení |
| Příklady organismů | Rostliny, živočichové, houby, prvoci | Bakterie, archaebakterie |
| Chloroplasty | Přítomny u rostlin a řas | Chybí |
Mikrofilamenta, známá také jako aktinová filamenta, jsou nejtenčí složkou cytoskeletu s průměrem přibližně sedm nanometrů. Skládají se z proteinu aktinu, který je jedním z nejhojnějších proteinů v eukaryotických buňkách. Aktinová filamenta mají zásadní význam pro buněčnou pohyblivost, zejména při tvorbě pseudopodií a při procesu zvaném améboidní pohyb. Tyto struktury jsou dynamické a mohou se rychle polymerizovat a depolymerizovat, což umožňuje buňce měnit svůj tvar a pohybovat se směrem k chemickým signálům nebo od nich pryč. Aktinová filamenta také spolupracují s motorickým proteinem myozinem při svalové kontrakci a při cytokinezi, tedy dělení cytoplazmy během buněčného dělení.
Intermediární filamenta představují střední kategorii cytoskeletálních struktur s průměrem asi deseti nanometrů. Na rozdíl od mikrofilament a mikrotubulů jsou intermediární filamenta stabilnější a méně dynamická. Jejich hlavní funkcí je poskytovat mechanickou odolnost buňkám proti napětí a deformaci. Existuje několik typů intermediárních filament, které jsou specifické pro různé typy buněk. Například keratinová filamenta nacházíme v epiteliálních buňkách, kde poskytují ochranu proti mechanickému poškození, zatímco neurofilamenta jsou charakteristická pro nervové buňky a podílejí se na udržování struktury axonů.
Mikrotubuly jsou nejsilnější složkou cytoskeletu s vnějším průměrem přibližně dvacet pět nanometrů. Jedná se o duté cylindrické struktury složené z proteinu tubulinu, konkrétně z dimerů alfa a beta tubulinu. Mikrotubuly hrají klíčovou roli v mnoha aspektech buněčné organizace a pohybu. Během buněčného dělení tvoří mitotické vřeténko, které zajišťuje správné rozdělení chromozomů do dceřiných buněk. Mikrotubuly také slouží jako dráhy pro intracelulární transport, po nichž se pohybují motorické proteiny kinesin a dynein nesoucí různé organely a vezikuly.
Buněčná pohyblivost je komplexní proces, který závisí na koordinované činnosti všech složek cytoskeletu. Eukaryotické buňky využívají několik mechanismů pohybu, z nichž každý je přizpůsoben specifickým podmínkám a funkcím. Améboidní pohyb, typický pro bílé krvinky a některé prvoky, je založen na tvorbě cytoplazmatických výběžků zvaných pseudopodie. Tento proces zahrnuje lokální polymerizaci aktinových filament na předním okraji buňky a jejich depolymerizaci vzadu, což vede k proudění cytoplazmy a pohybu celé buňky.
Řasinkový a bičíkový pohyb představuje další formu buněčné motility, která je charakteristická pro některé typy eukaryotických buněk. Řasinky a bičíky jsou výběžky buněčného povrchu obsahující strukturu zvanou axoném, která se skládá z devíti párů periferních mikrotubulů obklopujících dva centrální mikrotubuly. Motorický protein dynein způsobuje skluz mikrotubulů vůči sobě, což vede k ohýbání a vlnovému pohybu těchto struktur. Tato forma pohybu umožňuje pohyb celých buněk, jako jsou spermie, nebo transport tekutin a částic podél povrchu stacionárních buněk, například v dýchacích cestách.
Cytoskelet také umožňuje buňkám reagovat na vnější mechanické podněty a měnit svůj tvar v reakci na signály z okolního prostředí. Tento proces, známý jako mechanotransdukce, je důležitý pro mnoho biologických procesů včetně hojení ran, embryonálního vývoje a migrace buněk během imunitní odpovědi. Reorganizace cytoskeletu je řízena komplexní sítí signálních drah, které integrují informace z buněčného povrchu a koordinují odpovídající změny v architektuře cytoskeletu.
Buněčná membrána představuje základní strukturální a funkční prvek každé eukaryotické buňky, který odděluje vnitřní prostředí buňky od vnějšího okolí a zároveň umožňuje kontrolovanou výměnu látek a informací. Tato tenká, ale nesmírně důležitá struktura má tloušťku přibližně sedm až deset nanometrů a skládá se především z fosfolipidové dvojvrstvy, do které jsou zapuštěny různé proteiny a další molekuly.
Základní stavební jednotkou buněčné membrány jsou fosfolipidy, které mají charakteristickou amfipatickou povahu. To znamená, že jejich molekula obsahuje hydrofilní hlavičku směřující do vodného prostředí a dva hydrofobní ocasní řetězce orientované do vnitřku membrány. Díky této uspořádání vzniká stabilní dvojvrstva, která spontánně vytváří uzavřenou strukturu v přítomnosti vody. Membránové proteiny plní rozmanité funkce od transportu látek přes enzymatickou aktivitu až po přenos signálů do buňky.
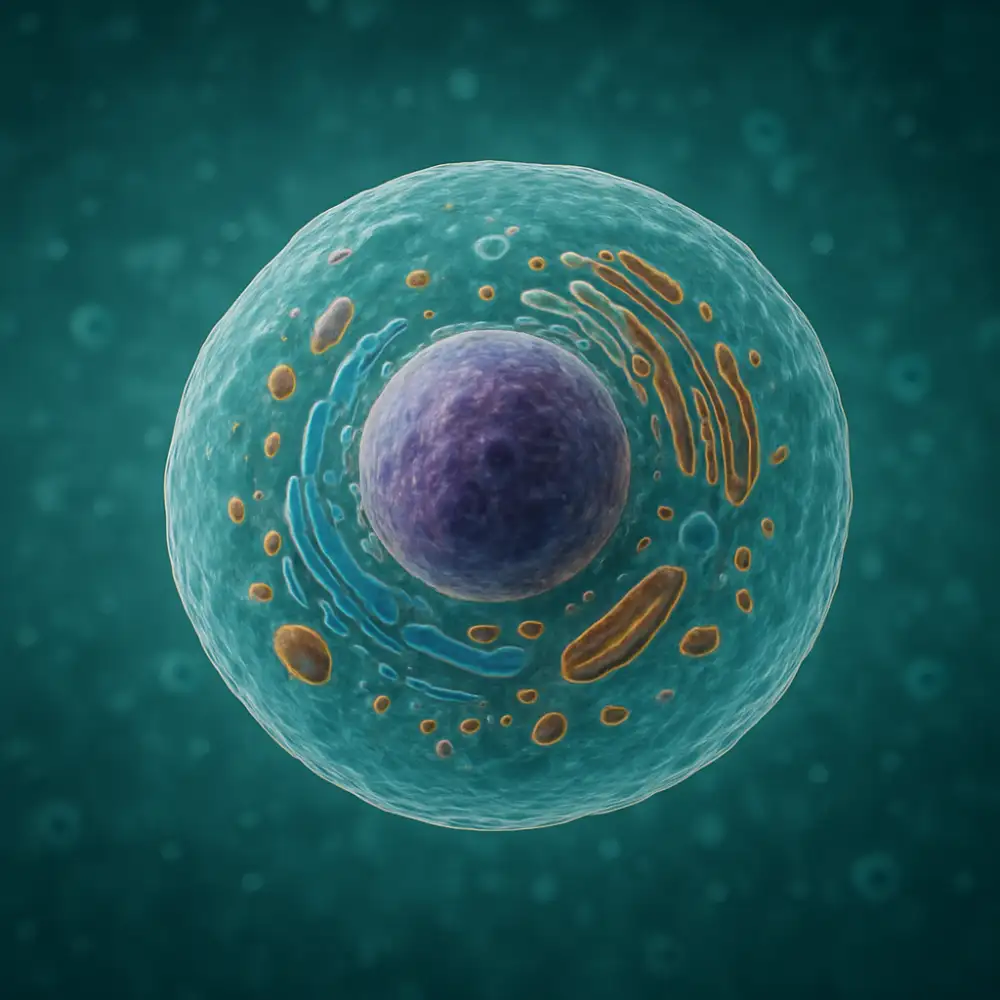
Selektivní propustnost představuje jednu z nejdůležitějších vlastností buněčné membrány. Membrána umožňuje volný průchod malým nepolárním molekulám jako je kyslík nebo oxid uhličitý, zatímco větší polární molekuly a ionty vyžadují specializované transportní mechanismy. Pasivní transport probíhá podle koncentračního gradientu bez spotřeby energie, přičemž může jít o jednoduchou difuzi nebo usnadněnou difuzi pomocí transportních proteinů. Aktivní transport naopak vyžaduje energii ve formě ATP a umožňuje přesun látek proti koncentračnímu gradientu, což je nezbytné pro udržení správného iontového složení buňky.
Receptorové proteiny v buněčné membráně zajišťují komunikaci buňky s vnějším prostředím. Tyto molekuly rozpoznávají specifické signální látky jako hormony nebo neurotransmitery a přenášejí informaci do nitra buňky, kde spouštějí kaskádu biochemických reakcí. Tento proces nazývaný buněčná signalizace je klíčový pro koordinaci funkcí v mnohobuněčných organismech.
Buněčná membrána také obsahuje různé sacharidové struktury, které jsou připojeny k lipidům nebo proteinům a tvoří glykokalyx na vnějším povrchu membrány. Tyto glykoproteiny a glykolipidy hrají důležitou roli v buněčném rozpoznávání, imunitní odpovědi a mezibuněčné komunikaci. Díky nim dokáží buňky rozlišovat mezi vlastními a cizími strukturami.
Cholesterol je další významnou složkou eukaryotických buněčných membrán, která ovlivňuje jejich fluiditu a stabilitu. Molekuly cholesterolu se vmezeřují mezi fosfolipidy a při vyšších teplotách snižují pohyblivost lipidových řetězců, zatímco při nižších teplotách brání jejich ztuhnutí. Tato regulace fluidity je nezbytná pro správnou funkci membránových proteinů.
Endocytóza a exocytóza představují mechanismy, kterými buňka přijímá nebo uvolňuje větší částice a makromolekuly. Při endocytóze se část membrány vchlípí dovnitř a vytvoří váček obsahující extracelulární materiál, zatímco při exocytóze se membránový váček z nitra buňky spojí s plazmatickou membránou a uvolní svůj obsah ven. Tyto procesy jsou zásadní pro výživu buňky, odstraňování odpadních látek a sekreci důležitých molekul.
Eukaryotická buňka představuje jeden z nejvýznamnějších evolučních skoků v historii života na Zemi, neboť její komplexní vnitřní struktura s membránově ohraničenými organelami umožnila vznik mnohobuněčných organismů a tím i celé rozmanitosti života, jak jej dnes známe
Radovan Šmída
Rostlinné a živočišné buňky představují dva základní typy eukaryotických buněk, které se vyvinuly odlišnými evolučními cestami a přizpůsobily se specifickým požadavkům svých organismů. Ačkoliv obě typy buněk sdílejí mnoho společných znaků typických pro eukaryoty, existují mezi nimi zásadní strukturální a funkční rozdíly, které odrážejí odlišné životní strategie rostlin a živočichů.
Nejviditelnějším rozdílem je přítomnost buněčné stěny u rostlinných buněk, která obklopuje plazmatickou membránu a poskytuje buňce mechanickou podporu a ochranu. Tato stěna je složena především z celulózy, hemicelulózy a pektinů, což jsou složité polysacharidy. Buněčná stěna umožňuje rostlinným buňkám odolávat vysokému osmotickému tlaku a udržovat svůj tvar. Naproti tomu živočišné buňky postrádají tuto rigidní strukturu a jsou ohraničeny pouze pružnou plazmatickou membránou, která jim umožňuje měnit tvar a pohybovat se, což je pro mnoho živočišných buněk zásadní vlastnost.
Dalším charakteristickým rysem rostlinných buněk je přítomnost chloroplastů, organelů odpovědných za fotosyntézu. Tyto dvojmembránové struktury obsahují chlorofyl a další pigmenty, které zachycují sluneční energii a přeměňují ji na chemickou energii ve formě glukózy. Chloroplasty jsou klíčové pro autotrofní výživu rostlin, která jim umožňuje vyrábět vlastní organické látky z anorganických zdrojů. Živočišné buňky chloroplasty neobsahují a jsou závislé na heterotrofní výživě, tedy na příjmu organických látek z vnějšího prostředí.
Vakuoly představují další významný rozdíl mezi těmito dvěma typy buněk. Rostlinné buňky typicky obsahují jednu velkou centrální vakuolu, která může zabírat až devadesát procent objemu buňky. Tato vakuola je naplněna buněčnou šťávou obsahující vodu, rozpuštené látky, pigmenty a různé metabolity. Vakuola slouží k udržování turgoru buňky, skladování živin a odpadních látek, a také k regulaci osmotické rovnováhy. Živočišné buňky mohou obsahovat menší vakuoly, ale ty jsou obvykle mnohem menší a slouží především k dočasnému skladování látek nebo k trávicím procesům.
Způsob dělení buněk se také liší mezi rostlinnými a živočišnými buňkami. Při cytokinezi, tedy rozdělení cytoplazmy, živočišné buňky vytváří brázdu, která postupně stahuje buňku uprostřed, dokud se nerozpůlí. Tento proces je řízen kontraktilním prstencem složeným z aktinových a myozinových filament. Rostlinné buňky však nemohou použít tento mechanismus kvůli přítomnosti rigidní buněčné stěny. Místo toho vytváří buněčnou destičku, která roste od středu buňky směrem k okrajům a postupně rozděluje mateřskou buňku na dvě dceřiné buňky.
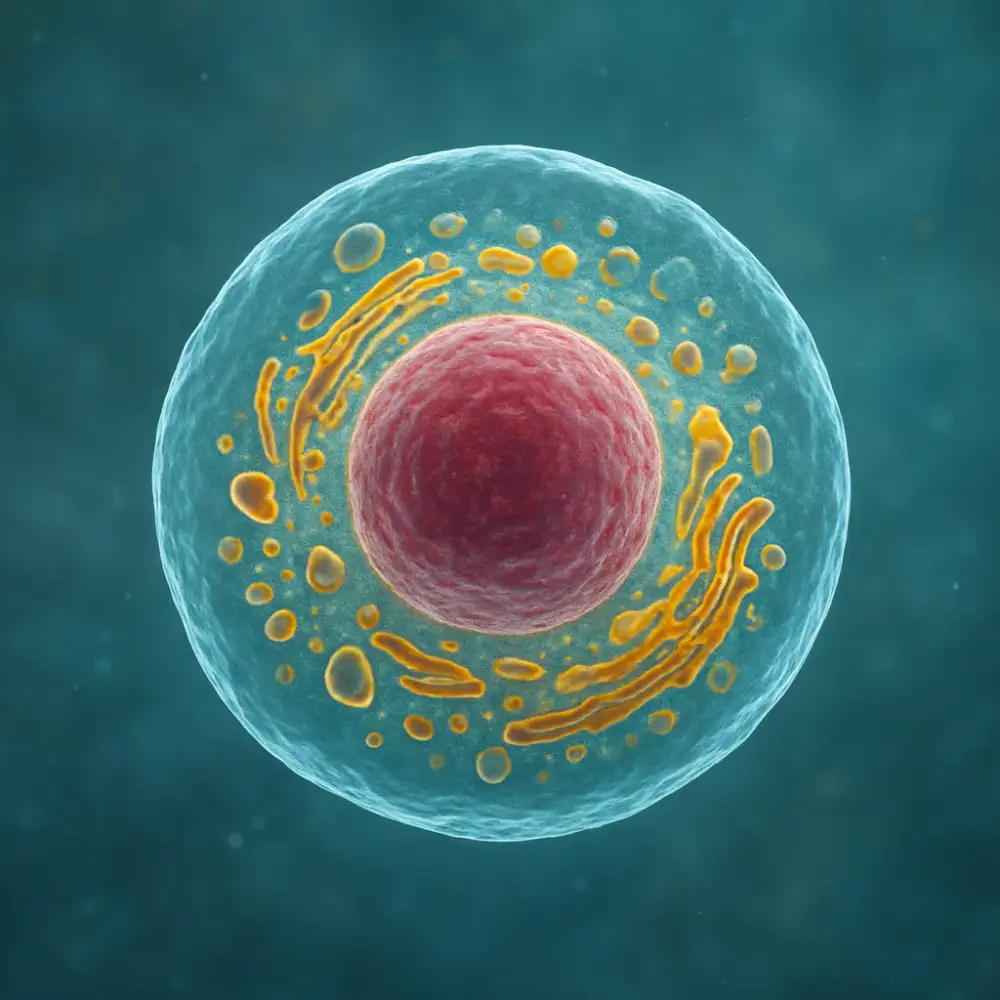
Centrosomy a centrioly jsou organely typické pro živočišné buňky, kde hrají důležitou roli při organizaci mikrotubulů během buněčného dělení. Většina rostlinných buněk tyto struktury postrádá, ačkoliv jsou schopny organizovat dělicí vřeténko pomocí alternativních mechanismů. Plasmodesmata jsou další unikátní strukturou rostlinných buněk, jedná se o cytoplazmatické můstky procházející buněčnou stěnou a spojující sousední buňky, což umožňuje přímou komunikaci a transport látek mezi buňkami. Živočišné buňky využívají k mezibuněčné komunikaci různé typy spojů, jako jsou těsné spoje, desmosomy nebo gap junctions, které mají odlišnou strukturu i funkci.
Buněčné dělení představuje jeden ze základních procesů života, který umožňuje růst, obnovu a reprodukci všech eukaryotických organismů. V přírodě existují dva hlavní typy buněčného dělení – mitóza a meióza, přičemž každý z nich má specifickou úlohu a průběh. Zatímco mitóza slouží k vytváření geneticky identických buněk pro růst a regeneraci tkání, meióza je specializovaný proces vedoucí ke vzniku pohlavních buněk s polovičním počtem chromozomů.
Mitóza je typ buněčného dělení, při kterém vznikají dvě geneticky identické dceřiné buňky se stejným počtem chromozomů jako měla původní mateřská buňka. Tento proces probíhá v somatických buňkách a je nezbytný pro růst organismu, hojení ran a obnovu opotřebovaných tkání. Před zahájením mitózy musí buňka projít interfází, během níž dochází k replikaci DNA a zdvojení genetického materiálu. Samotná mitóza se dělí na několik fází, které na sebe plynule navazují.
V profázi dochází ke kondenzaci chromatinu na viditelné chromozomy, které jsou tvořeny dvěma sesterskými chromatidami spojenými v oblasti centromery. Jaderná membrána se začína rozpadat a vytváří se mitotické vřeténko z mikrotubulů vycházejících z centriol. Prometafáze je charakterizována úplným rozpadem jaderné obálky a připojením chromozomů k vláknům mitotického vřeténka prostřednictvím kinetochorů. Následuje metafáze, kdy se všechny chromozomy uspořádají do roviny uprostřed buňky, což vytváří charakteristickou metafázovou destičku.
Anafáze představuje klíčový moment, kdy se sesterské chromatidy oddělují a jsou tahovány k opačným pólům buňky působením mikrotubulů vřeténka. V telofázi se chromozomy dekondenzují zpět na chromatin, kolem každé skupiny chromozomů se vytváří nová jaderná membrána a mitotické vřeténko se rozpadá. Celý proces je zakončen cytokinezí, při níž dochází k fyzickému rozdělení cytoplazmy a vzniku dvou samostatných dceřiných buněk.
Meióza je naproti tomu redukční dělení, které probíhá pouze při tvorbě pohlavních buněk – gamet. Tento proces je zásadní pro pohlavní rozmnožování, protože zajišťuje, že gamety obsahují pouze poloviční počet chromozomů. Při oplození se pak spojením dvou gamet obnovuje diploidní počet chromozomů typický pro daný druh. Meióza se skládá ze dvou po sobě jdoucích dělení – meiózy I a meiózy II, přičemž replikace DNA probíhá pouze jednou před první meiózou.
Meióza I je redukční dělení, při kterém se homologní chromozomy oddělují od sebe. V profázi I dochází k velmi důležitému procesu zvanému crossing-over neboli křížení, kdy si homologní chromozomy vyměňují úseky genetického materiálu. Tento proces je klíčový pro genetickou variabilitu potomstva. Profáze I je nejdelší a nejsložitější fází celé meiózy a dále se dělí na několik stadií. Během metafáze I se páry homologních chromozomů uspořádají do roviny buňky a v anafázi I se celé homologní chromozomy oddělují k opačným pólům, na rozdíl od mitózy, kde se oddělují sesterské chromatidy.
Po telofázi I a cytokinezi vznikají dvě haploidní buňky, které však stále obsahují chromozomy složené ze dvou sesterských chromatid. Následuje meióza II, která probíhá podobně jako mitóza. Během metafáze II se chromozomy opět uspořádají do roviny buňky a v anafázi II se konečně oddělují sesterské chromatidy. Výsledkem celého procesu meiózy jsou čtyři haploidní buňky s unikátní genetickou informací, které se mohou stát funkčními gametami.
Rozdíly mezi mitózou a meiózou jsou zásadní pro pochopení biologie eukaryotických organismů. Zatímco mitóza zachovává genetickou stabilitu a zajišťuje identické kopie buněk, meióza vytváří genetickou diverzitu prostřednictvím crossing-overu a náhodného rozdělení chromozomů. Tato variabilita je základem evoluce a adaptace druhů na měnící se prostředí.
Publikováno: 09. 11. 2025
Kategorie: Biologie