Genetický výzkum odhaluje unikátní starobylé odrůdy révy v etiopském Tigray
01. 08. 2026

Genetika jako vědecká disciplína má za sebou fascinující cestu plnou překvapivých objevů, omylů, geniálních intuicí a pečlivé laboratorní práce. Abychom pochopili, kde dnes stojíme, musíme se vrátit daleko do minulosti a sledovat, jak lidé postupně odhalovali tajemství dědičnosti.
Dlouho před tím, než vznikla genetika jako formální věda, lidé pozorovali, že potomci připomínají své rodiče. Zemědělci věděli, že křížením určitých odrůd plodin nebo plemen zvířat mohou dosáhnout žádoucích vlastností. Tato praktická znalost však postrádala teoretický základ. Neexistoval žádný systém, žádný zákon, který by vysvětloval, proč a jak se vlastnosti přenášejí z generace na generaci.
Skutečný průlom přišel v druhé polovině devatenáctého století díky práci augustiniánského mnicha Gregora Johanna Mendela. Mendel pracoval v Brně, kde prováděl systematické křížovací pokusy s hrachem setým. Jeho práce, publikovaná v roce 1866 pod názvem Versuche über Pflanzenhybriden, položila základy celé moderní genetiky. Mendel sledoval konkrétní znaky hrachu, jako byla barva semene, tvar lusku nebo výška rostliny, a pečlivě zaznamenával výsledky křížení v několika generacích. Z těchto pozorování odvodil dva základní zákony dědičnosti, které dnes nesou jeho jméno. Zákon o štěpení říká, že každý jedinec nese pro každý znak dvě vlohy, přičemž do pohlavních buněk přechází vždy jen jedna z nich. Zákon o volné kombinaci pak popisuje, jak se různé znaky kombinují nezávisle na sobě. Přestože Mendelova práce byla ve své době téměř zcela přehlížena, její znovuobjevení na počátku dvacátého století, kdy ji nezávisle na sobě znovu popsali Hugo de Vries, Carl Correns a Erich von Tschermak, znamenalo revoluci v biologickém myšlení.
Souběžně s tím se rozvíjela cytologie, věda o buňkách. Vědci jako Walther Flemming pozorovali pod mikroskopem podivné struktury v buněčném jádře, které se barvily speciálními barvivy. Tyto struktury dostaly název chromozomy. Flemming popsal v roce 1882 proces mitózy a ukázal, jak se chromozomy při buněčném dělení zdvojují a rozdělují do dceřiných buněk. Brzy nato byl objeven také proces meiózy, redukčního dělení, při němž vznikají pohlavní buňky s polovičním počtem chromozomů. Spojení Mendelových zákonů s pozorováním chromozomů vedlo ke vzniku chromozomové teorie dědičnosti, jejímiž hlavními autory byli Walter Sutton a Theodor Boveri. Tato teorie tvrdila, že Mendelovy vlohy jsou fyzicky umístěny na chromozomech.
Dalším klíčovým jménem v historii genetiky je Thomas Hunt Morgan, který pracoval s octomilkami rodu Drosophila. Morgan a jeho spolupracovníci prokázali, že geny jsou skutečně lokalizovány na chromozomech a že geny ležící na stejném chromozomu se dědí společně, což je jev nazývaný vazba genů. Morgan také popsal crossing-over, tedy výměnu úseků mezi homologními chromozomy při meióze, čímž vysvětlil, proč vazba genů není absolutní. Za svou práci obdržel v roce 1933 Nobelovu cenu za fyziologii a medicínu.
Přestože bylo jasné, že geny jsou na chromozomech, stále nebyla zodpovězena otázka, z čeho geny vlastně jsou. Chromozomy se skládají z bílkovin a z deoxyribonukleové kyseliny, zkráceně DNA. Dlouhou dobu se vědci domnívali, že nositelem genetické informace jsou bílkoviny, protože jsou strukturně rozmanitější. Zlom přišel v roce 1944, kdy Oswald Avery, Colin MacLeod a Maclyn McCarty prokázali na základě experimentů s bakteriemi, že genetická informace je přenášena právě DNA. Jejich práce navázala na dřívější experiment Fredericka Griffitha, který ukázal, že z mrtvých bakterií lze do živých přenést jakousi transformující substanci, která mění jejich vlastnosti.
Definitivní potvrzení přinesly v roce 1952 Alfred Hershey a Martha Chase, kteří pomocí radioaktivního značení prokázali, že při infekci bakteriofágem vstupuje do bakterie DNA, nikoli bílkovina. Cesta k odhalení struktury DNA tak byla otevřena. V roce 1953 publikovali James Watson a Francis Crick svůj slavný článek v časopise Nature, v němž popsali dvoušroubovicový model DNA. Tento model, vycházející mimo jiné z rentgenografických snímků Rosalind Franklinové a Mauricea Wilkinse, ukázal, že DNA se skládá ze dvou komplementárních řetězců stočených do šroubovice. Tato struktura okamžitě naznačovala, jak může být genetická informace kopírována a přenášena. Watson, Crick a Wilkins za tento objev obdrželi v roce 1962 Nobelovu cenu.
Navazující desetiletí přinesla objasnění genetického kódu, tedy způsobu, jakým sekvence nukleotidů v DNA určuje sekvenci aminokyselin v bílkovinách. Marshall Nirenberg, Har Gobind Khorana a Robert Holley rozluštili tento kód a ukázali, že každá aminokyselina je kódována trojicí nukleotidů, tzv. kodonem. Za tento objev získali Nobelovu cenu v roce 1968.
Sedmdesátá léta přinesla rozvoj rekombinantní DNA technologie. Herbert Boyer a Stanley Cohen ukázali, jak lze geny z různých organismů kombinovat a vkládat do bakterií, kde jsou exprimovány. Tato technologie otevřela dveře biotechnologiím a umožnila výrobu léků, jako je inzulin, pomocí geneticky modifikovaných bakterií. Následoval vývoj metody polymerázové řetězové reakce, PCR, kterou v roce 1983 vynalezl Kary Mullis. PCR umožňuje namnožit konkrétní úsek DNA milionkrát během několika hodin, což se stalo základním nástrojem moderní molekulární biologie, forenzní vědy i diagnostiky.

Vrcholem dvacátého století v oblasti genetiky byl bezesporu projekt lidského genomu, který byl zahájen v roce 1990 a jehož výsledky byly zveřejněny v roce 2003. Tento mezinárodní projekt přinesl kompletní sekvenci lidské DNA, čítající přibližně tři miliardy párů bází a obsahující odhadem dvacet až pětadvacet tisíc genů. Projekt lidského genomu otevřel zcela nové možnosti v medicíně, umožnil studium genetických základů nemocí a stal se základem pro rozvoj personalizované medicíny.
Dnes genetika stojí na prahu další revoluce díky technologii CRISPR-Cas9, která umožňuje cíleně editovat geny s dosud nevídanou přesností a dostupností. Tato metoda, za jejíž objev získaly Jennifer Doudna a Emmanuelle Charpentier Nobelovu cenu za chemii v roce 2020, slibuje nové způsoby léčby genetických chorob a otevírá zároveň závažné etické otázky. Pochopení celé této historické cesty je nezbytné pro každého, kdo chce genetice skutečně rozumět, a proto tvoří základ každé dobré genetiky učebnice.
Deoxyribonukleová kyselina, zkráceně DNA, představuje základní nositelku genetické informace ve všech živých organismech s výjimkou některých virů, které místo ní využívají RNA. Pochopení struktury DNA a mechanismů její replikace patří k absolutním základům moderní genetiky a bez těchto znalostí by nebylo možné porozumět ani dědičnosti, ani molekulárním procesům, které řídí život na buněčné úrovni.
Struktura DNA byla objasněna v roce 1953 Jamesem Watsonem a Francisem Crickem, kteří na základě rentgenových difrakčních snímků Rosalind Franklinové a Mauriceho Wilkinse navrhli model dvojité šroubovice. Tento model ukázal, že molekula DNA se skládá ze dvou polynukleotidových řetězců, které jsou vzájemně antiparalelně orientovány a stočeny kolem společné osy do tvaru pravotočivé šroubovice. Každý řetězec je tvořen opakujícími se jednotkami nazývanými nukleotidy, přičemž každý nukleotid obsahuje tři složky: deoxyribózu, fosfátovou skupinu a jednu ze čtyř dusíkatých bází.
Dusíkaté báze se dělí do dvou skupin. Puriny, adenin a guanin, mají dvouprstenčitou strukturu, zatímco pyrimidiny, cytosin a thymin, jsou jednoprstenčité. Klíčovým principem, který zajišťuje stabilitu dvojité šroubovice, je komplementarita bází. Adenin se vždy páruje s thyminem prostřednictvím dvou vodíkových vazeb, zatímco guanin se páruje s cytosinem pomocí tří vodíkových vazeb. Toto pravidlo, označované jako Chargaffovo pravidlo, bylo empiricky zjištěno ještě před navržením strukturního modelu a stalo se jedním z jeho klíčových potvrzení.
Fosfátové skupiny a deoxyribózové molekuly tvoří tzv. cukr-fosfátovou kostru, která tvoří vnější část šroubovice, zatímco párující se báze jsou orientovány dovnitř molekuly. Vzdálenost mezi sousedními páry bází je přibližně 0,34 nanometru a jeden úplný závit šroubovice odpovídá přibližně deseti párovým bazím, tedy délce asi 3,4 nanometru. Průměr celé molekuly je přibližně 2 nanometry.
V buňce není DNA přítomna jako volná molekula, ale je uspořádána do vyšších strukturních celků. U prokaryot se DNA nachází v oblasti nazývané nukleoid a je většinou organizována do jediné kruhové molekuly, která je superhelikálně stočena a asociována s různými proteiny. U eukaryot je situace podstatně složitější, protože DNA je uzavřena v membránou ohraničeném jádře a je organizována do chromozomů prostřednictvím hierarchického uspořádání. Na nejnižší úrovni se DNA navíjí kolem oktamerů histonových proteinů za vzniku nukleozomů, které připomínají korálky na niti. Tato struktura se dále kondenzuje do chromatinového vlákna o průměru 30 nanometrů, a v průběhu buněčného dělení dosahuje chromatin maximálního stupně kondenzace, který umožňuje spolehlivé rozdělení genetického materiálu do dceřiných buněk.
Replikace DNA je proces, při němž vznikají z jedné mateřské molekuly DNA dvě identické dceřiné molekuly. Tento proces je semikonzervativní, což znamená, že každá dceřiná molekula obsahuje jeden původní řetězec a jeden nově syntetizovaný řetězec. Tuto skutečnost experimentálně prokázali Matthew Meselson a Franklin Stahl v roce 1958 pomocí izotopového značení dusíku, přičemž jejich experiment je dodnes považován za jeden z nejelegantnějších v historii molekulární biologie.
Replikace začíná na specifických místech v molekule DNA, která se nazývají počátky replikace, anglicky origins of replication. U prokaryot existuje zpravidla jediný takový počátek, zatímco u eukaryot jich může být na jednom chromozomu tisíce, což umožňuje rychlou replikaci rozsáhlého genomu. V místě počátku replikace dochází k rozvinutí dvojité šroubovice a vzniku replikační vidličky, která se pohybuje oběma směry od počátku replikace.
Klíčovým enzymem replikace je DNA polymeráza, která katalyzuje syntézu nového řetězce přidáváním nukleotidů komplementárních k templátovému řetězci. DNA polymeráza však může syntetizovat nový řetězec pouze ve směru 5' na 3' a navíc potřebuje ke své činnosti krátký dvouřetězcový úsek, tedy tzv. primer. Primery jsou krátké RNA sekvence syntetizované enzymem primázou a po dokončení replikace jsou nahrazeny DNA.

Protože oba řetězce mateřské DNA jsou antiparalelní, musí být replikace jednoho z nich prováděna diskontinuálně. Vedoucí řetězec je syntetizován kontinuálně ve směru pohybu replikační vidličky, zatímco zpožděný řetězec je syntetizován po úsecích, které se nazývají Okazakiho fragmenty. Tyto fragmenty jsou po syntéze spojeny enzymem DNA ligázou do souvislého řetězce.
Celý proces replikace je doprovázen řadou kontrolních mechanismů, které zajišťují jeho přesnost. DNA polymeráza disponuje tzv. korektorskou aktivitou, díky níž dokáže rozpoznat a opravit chybně zařazené nukleotidy. Frekvence chyb při replikaci je díky těmto mechanismům extrémně nízká, přibližně jedna chyba na miliardu párů bází. Přesto mohou vznikat mutace, které jsou zdrojem genetické variability a hrají zásadní roli v evoluci.
Genetika jako věda se zrodila v okamžiku, kdy augustiniánský mnich Gregor Johann Mendel začal systematicky zkoumat dědičnost znaků u hrachu setého v zahradě brněnského kláštera. Jeho pokusy, prováděné v letech 1856 až 1863, přinesly výsledky, které tehdy téměř nikdo nedokázal ocenit. Teprve na počátku dvacátého století, kdy byly jeho práce znovuobjeveny nezávisle na sobě třemi vědci – Hugem de Vriesem, Carlem Corrensem a Erichem von Tschermak-Seysenegem – se ukázalo, že Mendel položil základy celé moderní genetiky.
| Parametr | Genetika (Jiří Šmarda, 2003) | Základy genetiky (Weiss a kol., 2016) | Lékařská genetika (Kočárek, 2008) | Genetika člověka (Strachan & Read, česky 2010) |
|---|---|---|---|---|
| Jazyk vydání | Čeština | Čeština | Čeština | Čeština (překlad) |
| Počet stran | 182 | 256 | 322 | 740 |
| Zaměření | Obecná genetika | Základy molekulární genetiky | Klinická a lékařská genetika | Humánní genetika |
| Cílová skupina | Studenti SŠ a VŠ | Studenti VŠ | Studenti lékařských fakult | Studenti VŠ a lékaři |
| Obtížnost | Středně pokročilá | Středně pokročilá | Pokročilá | Velmi pokročilá |
| Ilustrace a schémata | Ano, černobílé | Ano, barevné | Ano, barevné | Ano, barevné |
| Kapitoly o Mendelově genetice | Ano | Ano | Ano | Ano |
| Molekulární genetika | Základní přehled | Podrobně | Podrobně | Velmi podrobně |
| Genetické choroby | Okrajově | Částečně | Podrobně | Velmi podrobně |
| Přibližná cena (Kč) | 250 Kč | 390 Kč | 480 Kč | 1 200 Kč |
| Dostupnost | Antikvariáty | Knihkupectví a online | Knihkupectví a online | Knihkupectví a online |
| Cvičení a testy | Ano | Ano | Ano | Ano |
Mendel pracoval s hrachem setým (Pisum sativum) záměrně a promyšleně. Vybral si rostlinu, která se snadno pěstuje, má krátkou generační dobu a především vykazuje jasně rozlišitelné znaky, jako je barva semen, tvar semen, barva lusků, výška rostliny a mnoho dalších. Tyto znaky se navzájem nepřekrývaly a bylo možné je jednoznačně zařadit do jedné ze dvou kategorií. Právě tato volba experimentálního materiálu byla jedním z klíčových důvodů, proč Mendelovi jeho pokusy vyšly tak přesvědčivě.
První Mendelův zákon, zákon o uniformitě kříženců první generace, říká, že při křížení dvou homozygotních rodičovských linií lišících se v jednom znaku jsou všichni potomci první filiální generace (F1) uniformní, tedy fenotypově i genotypově stejní. Tento zákon vychází z předpokladu, že každý jedinec nese pro každý znak dvě alely, jednu od každého rodiče. Pokud je jeden rodič homozygotně dominantní (AA) a druhý homozygotně recesivní (aa), pak všichni potomci F1 budou heterozygoté (Aa) a budou vykazovat dominantní fenotyp. V učebnicích genetiky bývá tento zákon ilustrován klasickým příkladem křížení hrachu se žlutými a zelenými semeny, přičemž žlutá barva je dominantní nad zelenou.

Druhý Mendelův zákon, zákon o štěpení znaků, popisuje, co se děje v druhé filiální generaci (F2), tedy při křížení jedinců F1 mezi sebou. Heterozygotní jedinci Aa produkují gamety dvou typů – A a a – v poměru 1:1. Při náhodném spojení těchto gamet vznikají v generaci F2 jedinci v genotypovém poměru 1 AA : 2 Aa : 1 aa a ve fenotypovém poměru 3 dominantní : 1 recesivní. Tento poměr 3:1 je jedním z nejznámějších čísel v celé historii biologie a jeho správné pochopení je základním předpokladem pro zvládnutí dalšího studia genetiky.
Třetí Mendelův zákon, zákon o volné kombinovatelnosti znaků, jde ještě dál a sleduje dědičnost dvou různých znaků současně. Mendel zjistil, že alely pro různé znaky se do gamet rozdělují nezávisle na sobě, pokud leží na různých chromozomech. Při dihybridním křížení rodičů AABB × aabb vznikají v F2 potomci v poměru 9:3:3:1, kde devět jedinců vykazuje oba dominantní znaky, tři vykazují dominantní první a recesivní druhý znak, tři vykazují recesivní první a dominantní druhý znak a jeden jedinec je recesivní pro oba znaky. Tento zákon platí pouze tehdy, jsou-li sledované geny lokalizovány na různých chromozomech nebo jsou-li od sebe dostatečně vzdáleny na tomtéž chromozomu, aby mezi nimi docházelo k volné rekombinaci.
V moderních učebnicích genetiky jsou Mendelovy zákony prezentovány jako výchozí bod pro pochopení složitějších dědičných mechanismů. Studenti se učí, že realita je mnohdy komplikovanější než jednoduché mendelovské schéma. Existují jevy jako neúplná dominance, kdy heterozygot vykazuje fenotyp přechodný mezi oběma homozygoty, nebo kodominance, kdy se v fenotypu heterozygoта projevují oba alely současně a plně. Klasickým příkladem kodominance je krevní skupina AB u člověka, kdy červené krvinky nesou na svém povrchu jak antigen A, tak antigen B.
Dalším jevem, který rozšiřuje mendelovský pohled na dědičnost, je epistáze, tedy vzájemné ovlivňování alel různých genů. Pokud produkt jednoho genu potlačuje nebo modifikuje projev jiného genu, hovoříme o epistatickém vztahu. Tento jev vede k modifikovaným štěpným poměrům v F2 generaci, jako jsou poměry 9:7, 12:3:1 nebo 15:1, v závislosti na konkrétním typu epistáze.
Aplikace Mendelových zákonů v praxi je obrovská. V zemědělské genetice umožňují šlechtitelům předvídat výsledky křížení a cíleně vybírat kombinace znaků, které vedou k požadovaným vlastnostem nových odrůd. V lékařské genetice slouží mendelovské principy jako základ pro genetické poradenství, kdy odborníci pomáhají rodinám odhadnout pravděpodobnost, s níž se u jejich potomků může projevit dědičné onemocnění. Monogenně dědičná onemocnění, jako je cystická fibróza, srpkovitá anémie nebo Huntingtonova choroba, se řídí právě Mendlovými zákony a jejich dědičnost lze poměrně přesně předpovědět pomocí Punnetových čtverců nebo pravděpodobnostních výpočtů.
Punnetův čtverec, pojmenovaný po britském genetikovi Reginaldu Crundall Punnetovi, je grafická pomůcka, která usnadňuje výpočet pravděpodobnosti různých genotypových a fenotypových kombinací u potomků. V učebnicích genetiky je tato metoda vyučována jako jeden ze základních nástrojů genetické analýzy a studenti ji využívají při řešení genetických úloh. Přestože pro složitější případy, jako jsou polyhybridní křížení, se Punnetův čtverec stává nepraktickým, pro monohybridní a dihybridní křížení zůstává nezastupitelnou pomůckou.
Je důležité si uvědomit, že Mendelovy zákony nejsou dogmata, ale vědecké principy odvozené z experimentálních dat. Moderní genetika je doplňuje, upřesňuje a rozšiřuje, ale nikdy je nezpochybnila v jejich základní platnosti. Pochopení Mendelových zákonů dědičnosti je proto nezbytným předpokladem pro každého, kdo chce proniknout hlouběji do tajů genetiky, ať už jako student, vědec nebo lékař.
Každá živá buňka v těle nese v sobě pozoruhodnou strukturu, která uchovává veškeré dědičné informace organismu. Tato struktura, známá jako chromozom, představuje základní jednotku dědičnosti a hraje klíčovou roli ve všech procesech buněčného dělení. Chromozomy jsou tvořeny molekulami DNA těsně ovinutými kolem bílkovinných komplexů zvaných histony, přičemž celý tento komplex se nazývá chromatin. V době, kdy se buňka nepřipravuje na dělení, existuje chromatin v uvolněné, méně kondenzované formě, která umožňuje aktivní přepis genů a replikaci DNA.
Lidský organismus je charakteristický tím, že jeho somatické buňky obsahují 46 chromozomů uspořádaných do 23 párů. Tento stav označujeme jako diploidní, tedy 2n. Z těchto 23 párů tvoří 22 párů tzv. autosomy, tedy chromozomy nesoucí geny pro obecné tělesné znaky, a jeden pár tvoří pohlavní chromozomy, označované jako gonozomy. U žen jsou přítomny dva chromozomy X, zatímco u mužů je jeden chromozom X a jeden chromozom Y. Právě tento rozdíl v pohlavních chromozomech určuje genetické pohlaví jedince a ovlivňuje celou řadu znaků vázaných na pohlaví.
Buněčné dělení je proces, bez něhož by nebylo možné ani růst organismu, ani jeho reprodukce. Rozlišujeme dva základní typy buněčného dělení, a to mitózu a meiózu, přičemž každý z nich slouží jiným biologickým účelům a probíhá odlišným způsobem. Pochopení obou procesů je naprosto zásadní pro studium genetiky jako celku.
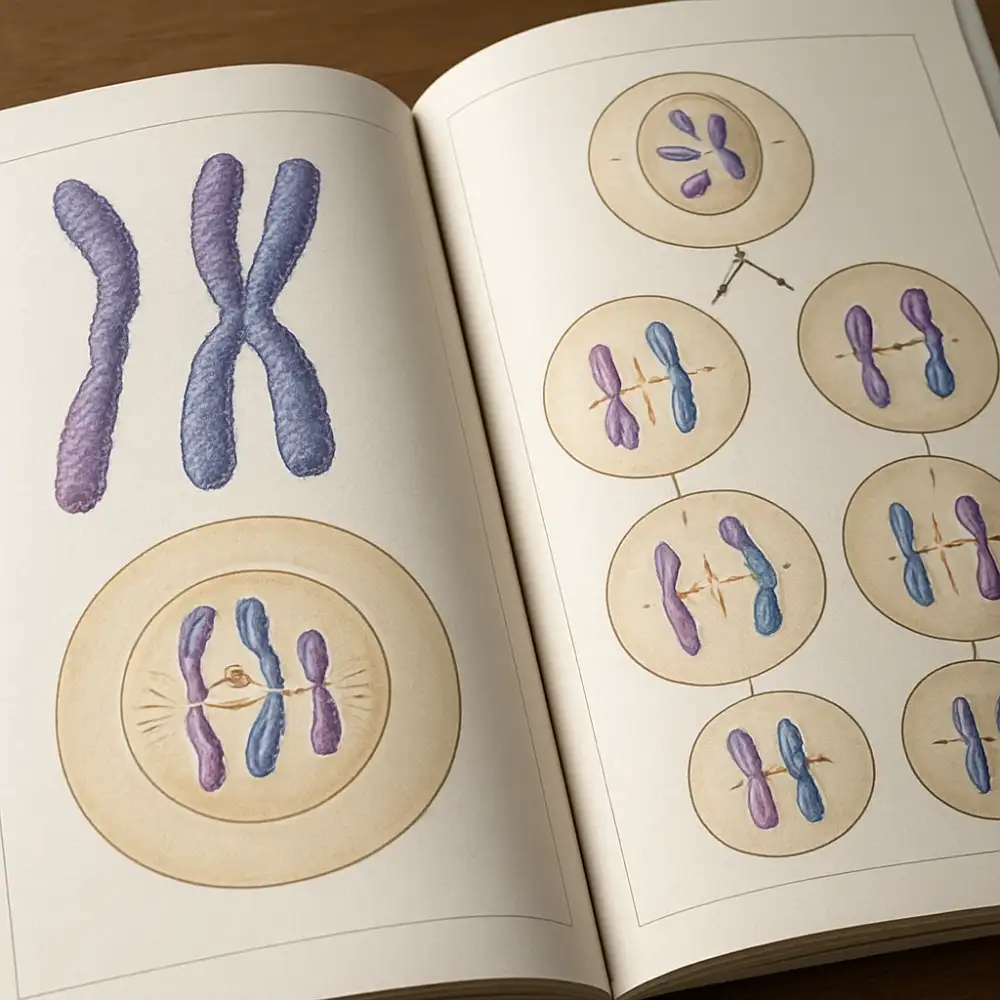
Mitóza je typ buněčného dělení, při němž z jedné mateřské buňky vznikají dvě dceřiné buňky se stejným počtem chromozomů, jaký měla buňka mateřská. Tento proces probíhá ve všech somatických buňkách a zajišťuje růst, opravu tkání a vegetativní rozmnožování. Mitóza se skládá z několika fází, které na sebe plynule navazují. Celý cyklus buněčného dělení začíná interfází, která sice není přímo součástí mitózy samotné, ale je pro ni naprosto nezbytná. V průběhu interfáze dochází k replikaci DNA, při níž se každý chromozom zdvojí a vznikají tzv. sesterské chromatidy spojené v místě zvaném centromera.
Po dokončení přípravné fáze nastupuje vlastní mitóza, která se tradičně dělí na čtyři hlavní fáze. První z nich je profáze, v níž chromozomy začínají kondenzovat a stávají se viditelné pod světelným mikroskopem. Jaderná membrána se začíná rozpadat a formuje se dělicí vřeténko tvořené mikrotubuly. Následuje metafáze, v níž jsou chromozomy maximálně kondenzovány a seřazeny v rovníkové rovině buňky, tzv. metafázové ploténce. Tato fáze je pro cytogenetiku obzvláště důležitá, protože právě v metafázi lze nejlépe studovat morfologii chromozomů a sestavovat karyotypy.
Ve třetí fázi, anafázi, dochází k oddělení sesterských chromatid, které jsou nyní nazývány dceřinými chromozomy. Tyto chromozomy jsou vlákny dělicího vřeténka taženy k opačným pólům buňky. Poslední fází je telofáze, při níž se chromozomy dekondenzují, obnovuje se jaderná membrána a buňka se fyzicky rozdělí procesem zvaným cytokineze. Výsledkem jsou dvě geneticky identické dceřiné buňky, každá s diploidním počtem chromozomů.
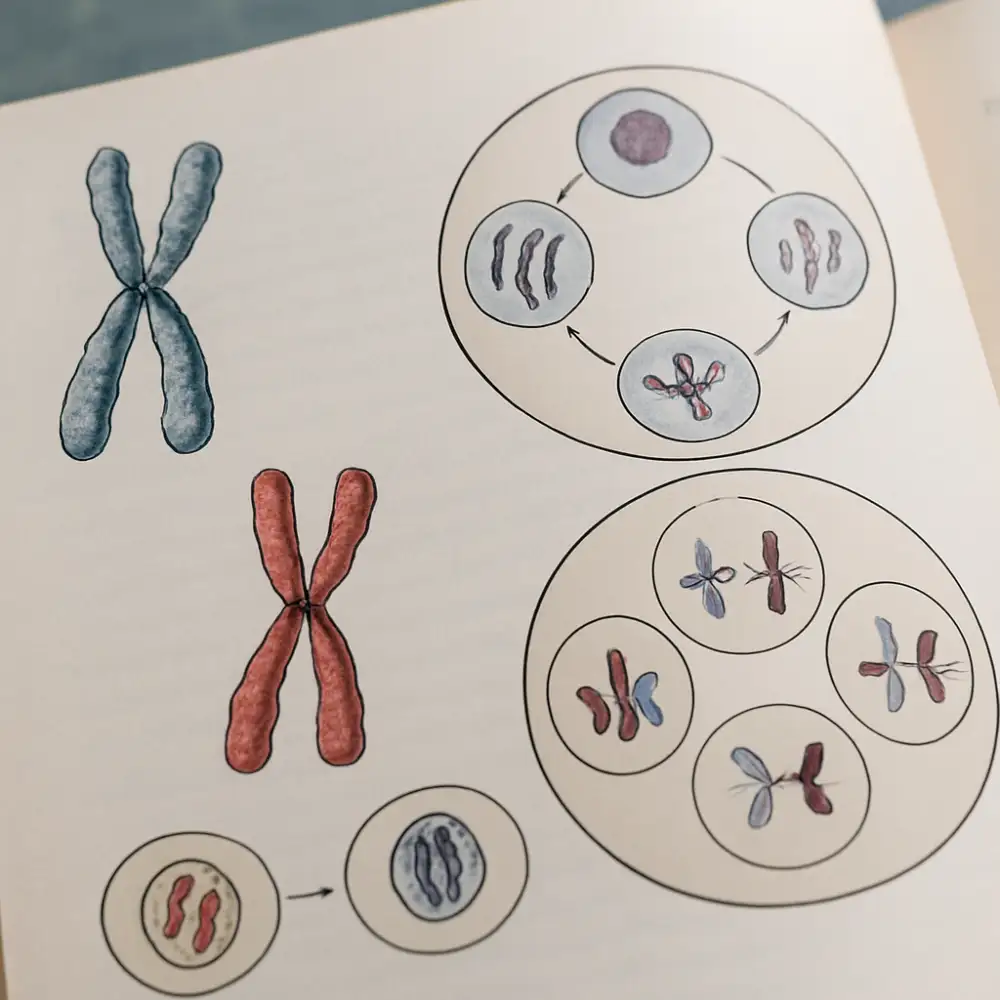
Zcela odlišný průběh má meióza, která je typická pro pohlavní buňky, tedy pro tvorbu gamet. Meióza je redukční dělení, při němž z jedné diploidní buňky vznikají čtyři haploidní buňky, každá s pouhými 23 chromozomy. Tento proces je nezbytný pro zachování správného počtu chromozomů při oplodnění, kdy splývají dvě haploidní gamety a obnovuje se diploidní stav. Meióza se skládá ze dvou po sobě jdoucích dělení, označovaných jako meióza I a meióza II.
Meióza I je redukční dělení, při němž dochází k oddělení homologních chromozomů. Její profáze je mimořádně složitá a dělí se na pět podfází: leptoten, zygoten, pachyten, diploten a diakinezi. Klíčovým dějem, který probíhá v průběhu profáze I, je crossing-over neboli překřížení. Při tomto procesu dochází k výměně úseků mezi homologními chromozomy, čímž vznikají nové kombinace alel. Crossing-over je jedním z nejvýznamnějších zdrojů genetické variability a má zásadní evoluční význam. K výměně genetického materiálu dochází na místech zvaných chiazmata, která jsou viditelná jako fyzická propojení mezi homologními chromozomy.
Po dokončení meiózy I, při níž vznikají dvě haploidní buňky s zdvojenými chromozomy, nastupuje meióza II, která svým průběhem připomíná mitózu. Dochází při ní k oddělení sesterských chromatid a výsledkem jsou čtyři haploidní buňky. U mužů z těchto čtyř buněk vznikají čtyři funkční spermie, zatímco u žen vzniká zpravidla pouze jedno funkční vajíčko a tři tzv. pólová tělíska, která zanikají.
Genetická rozmanitost, která vzniká v průběhu meiózy, je způsobena dvěma hlavními mechanismy. Prvním je již zmíněný crossing-over, druhým je náhodná segregace homologních chromozomů do dceřiných buněk. Vzhledem k tomu, že člověk má 23 párů homologních chromozomů, existuje teoreticky 2 na 23. mocninu možných kombinací, tedy více než 8 milionů různých kombinací chromozomů v jedné gametě, a to ještě bez uvažování crossing-overu. Tato čísla názorně ukazují, proč jsou geneticky identická sourozenci prakticky nemožná, s výjimkou jednovaječných dvojčat.
Poruchy v průběhu buněčného dělení mohou mít závažné genetické důsledky. Jednou z nejčastějších poruch je nondisjunkce, tedy nesprávné rozdělení chromozomů při dělení. Pokud k nondisjunkci dojde v průběhu meiózy, mohou vznikat gamety s nesprávným počtem chromozomů, což po oplodnění vede ke stavům označovaným jako aneuploidie. Nejznámějším příkladem aneuploidie u člověka je Downův syndrom, způsobený přítomností tří kopií chromozomu 21, tedy trizomií 21.
Genová exprese představuje jeden z nejzásadnějších procesů v celé biologii, protože právě skrze ni se genetická informace zakódovaná v molekule DNA stává funkčním produktem, který ovlivňuje stavbu a činnost každé živé buňky. Pochopení tohoto procesu je naprosto klíčové pro každého, kdo se chce orientovat v moderní genetice, a proto mu každá kvalitní genetika učebnice věnuje značnou pozornost.
Celý proces genové exprese se dělí do dvou hlavních etap – transkripce a translace. Tyto dvě fáze jsou na sebe navázány a spolupracují tak, aby z genetické informace vznikl konkrétní protein, který plní v organismu svoji specifickou roli. Bez dokonalého pochopení obou těchto kroků nelze plně porozumět tomu, jak geny ovládají vlastnosti živých organismů.
Transkripce je proces, při němž se genetická informace přepisuje z DNA do molekuly messenger RNA, tedy mRNA. Probíhá v buněčném jádře eukaryotických organismů, zatímco u prokaryot se odehrává přímo v cytoplazmě. Klíčovým enzymem celého procesu je RNA polymeráza, která se váže na specifickou oblast DNA nazývanou promotor. Promotor je sekvence nukleotidů, která signalizuje začátek genu a určuje, kde má polymeráza zahájit přepis. Po navázání na promotor enzym postupuje podél templátového vlákna DNA ve směru 3' na 5' a syntetizuje nové vlákno RNA ve směru 5' na 3'. Přitom se uplatňuje princip komplementarity bází, kdy adenin páruje s uracilem a guanin s cytosinem.
Transkripce se dělí do tří fází. Iniciace zahrnuje rozvinutí dvoušroubovice DNA a navázání RNA polymerázy na promotor. Poté nastupuje elongace, při níž polymeráza postupuje podél templátového vlákna a přidává ribonukleotidy k rostoucímu řetězci RNA. Celý proces je ukončen terminací, kdy polymeráza narazí na terminační sekvenci, přepis se zastaví a nově vzniklá molekula pre-mRNA se uvolní.
U eukaryot prochází primární transkript, tedy pre-mRNA, dalšími úpravami dříve, než opustí jádro. Jednou z nejdůležitějších úprav je sestřih, neboli splicing, při němž jsou z molekuly odstraněny nekódující úseky zvané introny a kódující úseky, exony, jsou navzájem spojeny. Tento proces zajišťuje komplex zvaný spliceosom. Kromě toho dochází k přidání čepičky na 5' konec molekuly a poly-A ocasu na konec 3'. Tyto modifikace chrání mRNA před degradací a usnadňují její export z jádra do cytoplazmy, kde probíhá druhá fáze genové exprese.
Translace je proces, při němž je informace zapsaná v sekvenci nukleotidů mRNA přeložena do sekvence aminokyselin tvořících polypeptidový řetězec. Probíhá na ribozomech, které jsou složeny z ribozomální RNA a proteinů. Ribozomy se skládají ze dvou podjednotek – velké a malé – které se spojují kolem molekuly mRNA a vytvářejí funkční komplex schopný syntézy proteinu.
Klíčovou roli při translaci hrají transferové RNA, tedy tRNA. Každá molekula tRNA nese na jednom konci specifický antikodon, trojici nukleotidů komplementárních ke kodonu mRNA, a na druhém konci příslušnou aminokyselinu. Genetický kód je tripletový, což znamená, že každé tři nukleotidy mRNA kódují jednu aminokyselinu. Celkem existuje 64 možných kodonů, přičemž 61 z nich kóduje aminokyseliny a tři jsou stop kodony signalizující ukončení syntézy.
Translace rovněž probíhá ve třech fázích. Během iniciace se malá podjednotka ribozomu váže na mRNA a hledá startovací kodon AUG, který kóduje methionin. Poté se připojí velká podjednotka a vzniká kompletní ribozom. V průběhu elongace se do ribozomu postupně přinášejí tRNA s příslušnými aminokyselinami, které jsou peptidovou vazbou spojovány do rostoucího řetězce. Ribozom se přitom pohybuje podél mRNA ve směru 5' na 3'. Terminace nastane, když ribozom narazí na stop kodon, k němuž se váže uvolňovací faktor, polypeptidový řetězec se oddělí a ribozom se rozpadne na své podjednotky.
Po uvolnění z ribozomu prochází nově syntetizovaný polypeptid dalšími úpravami, jako je skládání do trojrozměrné struktury, chemické modifikace nebo štěpení signálních sekvencí. Teprve po těchto posttranslačních modifikacích se protein stává plně funkčním a může plnit svoji biologickou roli.
Regulace genové exprese je stejně důležitá jako samotný průběh transkripce a translace. Buňka musí přesně řídit, které geny jsou v daném okamžiku aktivní, a to jak na úrovni transkripce prostřednictvím transkripčních faktorů a enhancerů, tak na úrovni translace nebo stability mRNA. Poruchy v regulaci genové exprese mohou vést k závažným onemocněním, včetně nádorových procesů.
Mutace představují jednu z nejzásadnějších kapitol celé genetiky, protože bez pochopení jejich podstaty, příčin a důsledků nelze plně uchopit mechanismy dědičnosti ani evoluci živých organismů. V každé kvalitní genetice učebnici najdeme mutace popsány jako změny v nukleotidové sekvenci DNA, které mohou být buď spontánní, nebo vyvolané vnějšími faktory. Tyto změny mohou postihovat jediný nukleotid, celé úseky chromozomu nebo dokonce celé sady chromozomů, a právě tato různorodost dělá z mutací téma neobyčejně komplexní a fascinující zároveň.
Z hlediska klasifikace rozlišujeme mutace na několika úrovních. Genové mutace, někdy nazývané také bodové mutace, jsou změny týkající se jednoho nebo několika nukleotidů v rámci jednoho genu. Patří sem substituce, při níž je jeden nukleotid nahrazen jiným, dále inzerce, kdy dojde k vložení jednoho či více nukleotidů do sekvence, a delece, při níž naopak nukleotid nebo více nukleotidů ze sekvence vypadne. Inzerce a delece jsou zvláště závažné, protože způsobují takzvaný posun čtecího rámce, což má za následek zásadní změnu aminokyselinové sekvence výsledného proteinu od místa mutace dál. Substituce se dělí na tranzice, kdy dochází k záměně purinu za purin nebo pyrimidinu za pyrimidin, a transverze, při nichž je purin nahrazen pyrimidinem nebo naopak.
Chromozomové mutace jsou závažnějšího rázu, protože postihují větší části chromozomů. Delece chromozomu znamená ztrátu části chromozomu, duplikace naopak zdvojení určitého úseku. Inverze je stav, kdy se část chromozomu otočí o 180 stupňů a vloží se zpět ve změněné orientaci, zatímco translokace označuje přesun části jednoho chromozomu na chromozom jiný. Tyto změny mohou mít dramatické důsledky pro fenotyp organismu a jsou spojeny s řadou vrozených vývojových vad.

Genomové mutace pak postihují celkový počet chromozomů v buňce. Polyploidie je stav, kdy organismus nese více než dvě sady chromozomů, a v rostlinné říši jde o jev poměrně běžný, který stál u zrodu mnoha kulturních plodin. Aneuploidie naopak označuje stav, kdy chybí jeden nebo více chromozomů, případně je jich přebývá. Nejznámějším příkladem aneuploidie u člověka je Downův syndrom, způsobený trizomií 21. chromozomu, tedy přítomností tří kopií tohoto chromozomu namísto dvou.
Příčiny mutací jsou rozmanité a genetika učebnice je tradičně rozděluje do dvou základních skupin. Spontánní mutace vznikají bez přičinění vnějších mutagenních faktorů, jsou důsledkem přirozených chyb při replikaci DNA nebo spontánní chemické nestability nukleotidových bází. Například tautomerní přesmyky, při nichž dochází k přechodné změně chemické formy bází, mohou způsobit nesprávné párování a následnou záměnu nukleotidu. Indukované mutace jsou naopak vyvolány působením mutagenů, tedy látek nebo fyzikálních faktorů, které zvyšují frekvenci mutací nad přirozenou míru.
Fyzikální mutageny zahrnují především ionizující záření, jako jsou rentgenové paprsky nebo záření gama, které mohou způsobovat zlomy v řetězcích DNA nebo vyvolávat tvorbu reaktivních kyslíkových radikálů poškozujících nukleotidy. Ultrafialové záření je typickým příkladem neionizujícího záření s mutagenním účinkem, přičemž způsobuje tvorbu pyrimidinových dimerů, zejména thymidínových dimerů, které brání správné replikaci DNA. Chemické mutageny jsou velmi různorodou skupinou zahrnující alkylační činidla, která přidávají alkylové skupiny k bazím a tím mění jejich párování, dále analogy bází, interkalační látky vkládající se mezi báze DNA a způsobující inzerce nebo delece, nebo deaminační látky měnící chemickou strukturu bází.
Biologické mutageny, jako jsou transpozony, představují zvláštní kategorii. Transpozony jsou pohyblivé genetické elementy schopné přemísťovat se v rámci genomu, a jejich vložení do funkčního genu může tento gen inaktivovat nebo narušit jeho regulaci. Retroviry mohou integrovat svou genetickou informaci do hostitelského genomu a tím způsobovat mutace na místě integrace.
Důsledky mutací jsou stejně pestré jako jejich příčiny a typy. Závisí na tom, ve které části genomu k mutaci dojde, zda postihuje kódující nebo nekódující oblast, zda jde o somatickou nebo zárodečnou buňku, a jaký je charakter samotné změny. Somatické mutace postihují pouze buňky těla a nejsou přenosné na potomstvo, mohou však vést ke vzniku nádorového bujení, pokud postihnou geny regulující buněčný cyklus. Zárodečné mutace jsou přenášeny na potomstvo a mohou být příčinou dědičných onemocnění.
Z hlediska dopadu na funkci proteinu rozlišujeme mutace tiché, neboli synonymní, při nichž dojde ke změně kodonu, ale výsledná aminokyselina zůstane stejná díky degeneraci genetického kódu. Missense mutace vedou k záměně jedné aminokyseliny za jinou, přičemž důsledky závisí na tom, jak zásadní roli postižená aminokyselina hraje ve struktuře a funkci proteinu. Nonsense mutace jsou zvláště závažné, protože mění smysluplný kodon na stop kodon a způsobují předčasné ukončení translace, což zpravidla vede ke vzniku nefunkčního proteinu.
Je důležité si uvědomit, že ne každá mutace je škodlivá. Evoluce je z velké části poháněna právě mutacemi, které přinášejí novou genetickou variabilitu, na níž pak může působit přirozený výběr. Většina mutací je neutrálních nebo mírně škodlivých, jen malá část je výrazně škodlivá a ještě menší část může být za určitých podmínek výhodná. Tato komplexní sohra mutací, selekce a genetického driftu tvoří základ moderního chápání evoluce, jak je popsáno v každé moderní genetice učebnici.
Populační genetika představuje fascinující odvětví biologie, které se zabývá studiem genetické variability v rámci celých populací organismů, nikoli pouze jednotlivých jedinců. Zatímco klasická mendelovská genetika nás učí, jak se znaky dědí z rodičů na potomky, populační genetika rozšiřuje tento pohled na mnohem širší měřítko a zkoumá, jak se frekvence alel mění v čase a prostoru napříč celými skupinami jedinců téhož druhu.
Základním pilířem populační genetiky je Hardy-Weinbergův zákon, který byl nezávisle na sobě formulován britským matematikem Godfreym Haroldem Hardym a německým lékařem Wilhelmem Weinbergem v roce 1908. Tento zákon, někdy nazývaný také Hardy-Weinbergova rovnováha nebo Hardy-Weinbergův princip, popisuje podmínky, za nichž zůstávají frekvence alel a genotypů v populaci konstantní z generace na generaci. Je to vlastně matematický model ideální populace, který slouží jako referenční bod pro studium skutečných populací v přírodě.
Aby platila Hardy-Weinbergova rovnováha, musí být splněno několik předpokladů. Populace musí být nekonečně velká, nebo alespoň dostatečně velká, aby nedocházelo k náhodným výkyvům frekvencí alel způsobeným genetickým driftem. Dále musí probíhat náhodné párování jedinců, tedy panmixie, bez jakýchkoli preferencí při výběru partnera. Nesmí docházet k mutacím, které by měnily frekvence alel, ani k migraci jedinců do populace nebo z ní. A konečně, přirozený výběr nesmí upřednostňovat žádný z genotypů, tedy všechny genotypy musí mít stejnou reprodukční zdatnost, neboli fitness.

Matematické vyjádření Hardy-Weinbergova zákona je elegantně jednoduché. Pokud označíme frekvenci jedné alely písmenem p a frekvenci druhé alely písmenem q, přičemž platí, že p + q = 1, pak frekvence jednotlivých genotypů v rovnovážné populaci jsou dány vztahem p² + 2pq + q² = 1. Homozygoti pro dominantní alelu mají frekvenci p², heterozygoti mají frekvenci 2pq a homozygoti pro recesivní alelu mají frekvenci q². Tento jednoduchý vzorec má obrovský praktický význam, protože nám umožňuje odhadovat frekvence alel v populaci na základě pozorované frekvence homozygotních recesivních jedinců, kteří jsou fenotypově rozpoznatelní.
Vezměme si konkrétní příklad. Pokud víme, že v určité populaci se recesivní onemocnění, jako je například cystická fibróza, vyskytuje u jednoho ze dvou tisíc pět set jedinců, pak frekvence homozygotů pro recesivní alelu je q² = 1/2500, tedy q² = 0,0004. Z toho vyplývá, že q = 0,02, tedy frekvence recesivní alely v populaci je dvě procenta. Frekvence dominantní alely pak bude p = 1 - 0,02 = 0,98. Frekvence přenašečů, tedy heterozygotů, bude 2pq = 2 × 0,98 × 0,02 = přibližně 0,039, což znamená, že přibližně každý dvacátý pátý jedinec v populaci je přenašečem tohoto onemocnění, aniž by o tom věděl.
Skutečné populace se od ideálního Hardy-Weinbergova modelu více či méně odchylují, a právě tyto odchylky jsou pro populační genetiky nesmírně zajímavé. Genetický drift, neboli náhodný genetický posun, je jev, který se projevuje zejména v malých populacích. Jde o náhodné změny frekvencí alel způsobené tím, že do další generace se přenáší pouze část alel z generace předchozí, a to nikoli vždy v přesném poměru odpovídajícím frekvencím v rodičovské generaci. V extrémních případech může genetický drift vést k úplné fixaci jedné alely, kdy všichni jedinci v populaci jsou homozygotní pro tuto alelu, nebo naopak k úplné ztrátě alely z populace.
Zvláštním případem genetického driftu je efekt hrdla láhve, anglicky bottleneck effect, kdy dojde k dramatickému poklesu velikosti populace v důsledku nějaké katastrofické události, jako je epidemie, přírodní katastrofa nebo nadměrný lov. Přeživší jedinci představují pouze malý a náhodný vzorek původní genetické variability, a proto může být genetická diverzita nové populace výrazně snížena. Podobným jevem je efekt zakladatele, kdy novou populaci zakládá malý počet jedinců, kteří se oddělili od původní populace a osídlili nové území. Genetická variabilita takové populace je omezena na alely přítomné u těchto zakladatelů.
Přirozený výběr je dalším mocným faktorem, který narušuje Hardy-Weinbergovu rovnováhu. Pokud jsou jedinci s určitým genotypem úspěšnější v přežívání a rozmnožování než jedinci s jinými genotypy, frekvence příslušných alel se bude v populaci postupně měnit. Selekce může působit různými způsoby – může upřednostňovat homozygoty pro jednu z alel, může zvýhodňovat heterozygoty, nebo může naopak heterozygoty znevýhodňovat. Příkladem výhodnosti heterozygotů je srpkovitá anémie, kde heterozygoti jsou v oblastech výskytu malárie odolnější vůči tomuto onemocnění než homozygoti pro normální hemoglobin, přičemž homozygoti pro mutantní alelu trpí závažnou formou srpkovité anémie.
Migrace, neboli tok genů mezi populacemi, představuje další faktor ovlivňující genetickou strukturu populací. Příchod nových jedinců z jiných populací přináší do místní populace nové alely nebo mění frekvence již přítomných alel. Tok genů má obecně tendenci snižovat genetické rozdíly mezi populacemi a zvyšovat genetickou variabilitu uvnitř každé z nich.
Mutace jsou sice základním zdrojem genetické variability, ale jejich přímý vliv na změny frekvencí alel v krátkodobém měřítku je obvykle velmi malý, protože mutace jsou vzácné jevy. Přesto jsou v dlouhodobém evolučním pohledu nenahraditelné, protože bez mutací by neexistoval žádný zdroj nových alel, na které by mohl působit přirozený výběr.
Praktické aplikace Hardy-Weinbergova zákona jsou velmi rozsáhlé. V lékařské genetice se využívá pro odhad frekvencí přenašečů genetických onemocnění v populaci, což je důležité pro genetické poradenství a plánování preventivních programů. Ve forenzní genetice se Hardy-Weinbergovy frekvence používají při výpočtu pravděpodobnosti shody DNA profilů. V evoluční biologii slouží jako základ pro studium evolučních procesů a jejich vlivu na genetickou strukturu populací.
Je důležité si uvědomit, že Hardy-Weinbergův zákon není pouze abstraktní matematický konstrukt, ale mocný nástroj, který nám pomáhá pochopit, jak genetická variabilita vzniká, udržuje se a mění v průběhu evoluce. Porovnáváním pozorovaných frekvencí genotypů s frekvencemi očekávanými podle Hardy-Weinbergovy rovnováhy mohou vědci odhalovat, které evoluční síly působí na danou populaci a jak intenzivně. Tato schopnost detekovat odchylky od rovnováhy a interpretovat je v kontextu evolučních mechanismů dělá z Hardy-Weinbergova zákona jeden z nejdůležitějších konceptů celé moderní genetiky a evoluční biologie.

Molekulární genetika představuje jedno z nejdynamičtěji se rozvíjejících odvětví moderní biologie, přičemž metody, které dnes výzkumníci využívají, by ještě před několika desetiletími působily jako vědecká fantastika. Pochopení těchto metod je klíčové nejen pro studenty biologie a medicíny, ale i pro každého, kdo chce skutečně porozumět tomu, jak funguje dědičnost na té nejzákladnější úrovni – na úrovni molekul DNA.
Jednou z nejrevolucionářštějších technik, která zcela proměnila způsob, jakým vědci pracují s genetickým materiálem, je polymerázová řetězová reakce, obecně známá pod zkratkou PCR. Tato metoda, vyvinutá Kary Mullisem v roce 1983, umožňuje namnožit specifický úsek DNA do milionů kopií během několika hodin. Princip spočívá v opakovaném cyklování teplot, přičemž v každém cyklu dochází k denaturaci dvoušroubovice, nasedání primerů a syntéze nových vláken pomocí termostabilní DNA polymerázy. Bez PCR by dnes nebylo možné provádět diagnostiku infekčních onemocnění, forenzní analýzy ani výzkum genetických mutací spojených s nádorovými onemocněními.
Dalším zásadním nástrojem moderní molekulární genetiky je sekvenování DNA. Sangerova metoda, která dominovala výzkumu po několik desetiletí, byla postupně nahrazena takzvanými metodami sekvenování nové generace, anglicky označovanými jako NGS – Next Generation Sequencing. Tyto přístupy umožňují paralelní sekvenování obrovského množství úseků DNA současně, čímž dramaticky snižují náklady i čas potřebný k přečtení celého genomu. Dnes je možné osekvenovat celý lidský genom během jediného dne, zatímco projekt Lidský genom, který byl dokončen v roce 2003, trval přes deset let a stál miliardy dolarů. Tato technologická revoluce otevřela dveře personalizované medicíně, kde je léčba přizpůsobena konkrétnímu genetickému profilu pacienta.
Nesmíme opomenout ani technologii CRISPR-Cas9, která je v současnosti pravděpodobně nejdiskutovanější metodou v celé genetice. Tento systém, původně objevený jako součást imunitní obrany bakterií, byl adaptován pro přesné editování genomu prakticky jakéhokoli organismu. Pomocí CRISPR-Cas9 je možné vystřihnout konkrétní gen, opravit mutaci nebo vložit novou sekvenci na přesně definované místo v genomu. Nobelova cena za chemii v roce 2020, udělená Jennifer Doudna a Emmanuelle Charpentier, potvrdila, jak zásadní tento objev pro vědu je. V kontextu genetiky jako vědecké disciplíny i jako učebnicového předmětu představuje CRISPR přelomový moment, který bude ovlivňovat medicínu, zemědělství i etické debaty ještě po mnoho generací.
Hybridizační techniky, jako je fluorescenční in situ hybridizace označovaná jako FISH, umožňují vizualizovat specifické sekvence DNA přímo na chromozomech pomocí fluorescenčně značených sond. Tato metoda nachází uplatnění zejména v cytogenetice, kde pomáhá odhalit chromozomální přestavby, delece nebo amplifikace genů spojené s různými genetickými syndromy nebo nádorovými onemocněními. Kombinace klasické cytogenetiky s molekulárními přístupy tak vytváří mocný diagnostický nástroj.
Analýza genové exprese pomocí mikročipů a RNA sekvenování, označovaného jako RNA-seq, umožňuje vědcům sledovat, které geny jsou aktivní v konkrétním typu buňky, v konkrétním vývojovém stádiu nebo za určitých podmínek prostředí. Tím se otvírá zcela nový pohled na regulaci genů a na to, jak se z jednoho genomu mohou vyvinout stovky různých typů buněk se zcela odlišnými funkcemi. Transkriptomika, jak se tato oblast nazývá, se stala nepostradatelnou součástí výzkumu rakoviny, vývojové biologie i neurověd.
Proteomika a genomika spolu úzce spolupracují v rámci systémové biologie, která se snaží porozumět živým organismům jako celku, nikoliv pouze jako součtu jednotlivých genů. Hmotnostní spektrometrie umožňuje identifikovat a kvantifikovat tisíce proteinů najednou, čímž doplňuje informace získané na úrovni DNA a RNA o funkční rozměr. Teprve tehdy, když víme, jaké proteiny buňka skutečně produkuje a v jakém množství, můžeme plně pochopit, jak se genetická informace projevuje v živém organismu.
Epigenetika, která studuje dědičné změny v genové expresi bez změny samotné sekvence DNA, přinesla do molekulární genetiky zcela nový pohled. Metylace DNA a modifikace histonů jsou mechanismy, které ovlivňují, zda bude určitý gen aktivní nebo umlčený, přičemž tyto vzorce mohou být přenášeny z buňky na buňku i z generace na generaci. Bisulfitové sekvenování je jednou z klíčových metod, která umožňuje mapovat metylaci DNA v celogenomovém měřítku a odhalovat tak epigenetické změny spojené s vývojem, stárnutím nebo nemocí.
Moderní molekulárně genetické metody tak tvoří komplexní soubor nástrojů, které navzájem doplňují a prohlubují naše poznání genetiky. Každá z těchto technik přináší jiný pohled na genetickou informaci – od její struktury přes expresi až po funkci – a teprve jejich kombinace nám umožňuje skutečně porozumět složitosti života na molekulární úrovni.
Genetické inženýrství představuje jednu z nejrevolučnějších disciplín moderní biologie, která zásadně proměnila nejen vědecký výzkum, ale také každodenní život lidí po celém světě. V rámci studia genetiky, jak ji zachycují učebnice určené středoškolským i vysokoškolským studentům, tvoří tato oblast stále důležitější součást osnov, protože pochopení základních principů manipulace s DNA je dnes považováno za nezbytnou součást biologické gramotnosti.

Základním nástrojem genetického inženýrství jsou restrikční endonukleázy, tedy enzymy schopné rozpoznat specifické sekvence DNA a přestřihnout ji na přesně definovaných místech. Tyto enzymy byly původně objeveny jako součást bakteriálního obranného systému proti virům, avšak vědci záhy pochopili jejich obrovský biotechnologický potenciál. Díky nim je možné izolovat konkrétní gen z genomu jednoho organismu a vložit jej do genomu organismu jiného, čímž vznikají takzvané rekombinantní organismy. Tento postup stojí za výrobou celé řady léčiv, která dnes zachraňují miliony životů.
Jedním z nejznámějších příkladů praktického využití genetického inženýrství je výroba inzulinu. Ještě před několika desetiletími byl inzulin pro diabetiky získáván z pankreatu prasat nebo skotu, což s sebou přinášelo řadu komplikací včetně alergických reakcí. Dnes je lidský inzulin produkován bakteriemi Escherichia coli nebo kvasinkami, do jejichž genomu byl vložen gen kódující tuto bílkovinu. Výsledný produkt je identický s lidským inzulinem, je dostupnější a bezpečnější. Právě tento příklad bývá v učebnicích genetiky uváděn jako modelový případ úspěšné aplikace rekombinantní DNA technologie.
Dalším klíčovým pojmem, který studenti genetiky musí dobře pochopit, jsou vektory – přenašeče genetické informace. Nejčastěji se využívají plazmidy, což jsou malé kruhové molekuly DNA přirozeně se vyskytující v bakteriích. Plazmid je možné v laboratoři otevřít pomocí restrikčních enzymů, vložit do něj žádaný gen a poté jej zavést zpět do bakteriální buňky. Bakterie pak tento gen čte a produkuje odpovídající protein. Vedle plazmidů se jako vektory využívají také virové částice, zejména v případech, kdy je třeba dodat genetickou informaci do buněk savců, například při genové terapii.
Genová terapie představuje oblast, která vzbuzuje velká očekávání i etické diskuse zároveň. Principiálně jde o opravu nebo náhradu nefunkčního genu v buňkách pacienta. Některé formy genové terapie jsou již klinicky schváleny a používají se například při léčbě určitých typů dědičné slepoty nebo spinální muskulární atrofie. Přestože jde o relativně mladou disciplínu, její pokrok je mimořádně rychlý a učebnice genetiky musí být v tomto ohledu neustále aktualizovány, aby odrážely nejnovější poznatky.
Biotechnologie v zemědělství jsou dalším rozsáhlým polem, kde se genetické inženýrství uplatňuje v masovém měřítku. Geneticky modifikované organismy, zkráceně GMO, jsou rostliny nebo živočichové, jejichž genetická výbava byla záměrně pozměněna za účelem dosažení určitých vlastností. Nejrozšířenějšími příklady jsou kukuřice nebo sója odolné vůči herbicidům nebo hmyzím škůdcům. Tyto plodiny jsou schopny produkovat vlastní insekticidní protein díky genu převzatému z bakterie Bacillus thuringiensis, což výrazně snižuje potřebu chemických postřiků. Kritici však upozorňují na možné ekologické dopady, jako je šíření modifikovaných genů do volné přírody nebo vznik rezistentních populací škůdců.
Moderní genetika přinesla také technologii CRISPR-Cas9, která je v současných učebnicích genetiky popisována jako skutečná revoluce v přesném editování genomu. Na rozdíl od dřívějších metod umožňuje CRISPR cílit na konkrétní sekvenci DNA s nebývalou přesností a efektivitou. Využití nachází jak ve výzkumu základních biologických procesů, tak v medicíně, zemědělství i průmyslu. Pomocí této technologie již bylo možné vytvořit prasata s genomem upraveným tak, aby jejich orgány byly vhodné pro transplantaci do lidského těla, nebo rostliny s vyšším nutričním obsahem.
Etické otázky spojené s genetickým inženýrstvím jsou nedílnou součástí výuky genetiky a v dobrých učebnicích jim bývá věnována samostatná kapitola. Společnost musí zvažovat, kde leží hranice přijatelného zásahu do přirozeného genomu, kdo nese odpovědnost za případné negativní důsledky a jak zajistit rovný přístup k biotechnologickým inovacím. Tyto otázky nemají jednoznačné odpovědi, ale jejich promýšlení je nezbytné pro každého, kdo chce genetice skutečně rozumět nejen jako vědě, ale také jako součásti lidské civilizace.
Genetika není jen věda o dědičnosti, je to příběh života samotného, zapsaný v jazyce molekul, který čteme stránku po stránce jako učebnici vesmíru – každý gen je kapitolou, každá mutace obratem děje a každý organismus živým důkazem toho, že evoluce píše své romány bez konce.
Radovan Šimánek
Epigenetika představuje fascinující oblast moderní genetiky, která zásadním způsobem rozšiřuje naše chápání dědičnosti. Po desetiletí bylo v učebnicích genetiky prezentováno dogma, podle něhož jsou dědičné vlastnosti organismu určeny výhradně sekvencí nukleotidů v molekule DNA. Tento pohled byl sice správný, avšak zdaleka ne úplný. Epigenetika se zabývá studiem dědičných změn v genové expresi, které nejsou způsobeny změnami v samotné sekvenci DNA, ale přesto mohou být předávány z buňky na buňku, a v některých případech dokonce z generace na generaci.

Abychom pochopili podstatu epigenetických mechanismů, je třeba si uvědomit, jak je DNA v buňce uspořádána. DNA není v jádře buňky volně rozptýlena, nýbrž je těsně navinuta kolem speciálních bílkovinných komplexů zvaných nukleozomy, které jsou tvořeny proteiny označovanými jako histony. Toto uspořádání umožňuje, aby se do jaderného prostoru vešly metry dlouhé molekuly DNA. Způsob, jakým je DNA kolem histonů navinuta, a chemické modifikace, které tyto histony nesou, mají přímý vliv na to, které geny jsou aktivní a které jsou umlčeny.
Jedním z nejlépe prostudovaných epigenetických mechanismů je metylace DNA. Při tomto procesu dochází k přidání metylové skupiny na cytozinové báze v DNA, nejčastěji v místech, kde se cytozin nachází bezprostředně před guaninem, tedy v takzvaných CpG dinukleotidech. Metylace promotorových oblastí genů zpravidla vede k jejich umlčení, protože metylované oblasti jsou hůře přístupné pro transkripční faktory a RNA polymerázu. Tento mechanismus hraje klíčovou roli v procesech diferenciace buněk, kdy se z jedné oplodněné buňky vyvíjejí stovky různých buněčných typů, přestože všechny nesou totožnou genetickou informaci.
Druhým zásadním epigenetickým mechanismem jsou modifikace histonů. Histony mohou být chemicky modifikovány na svých volných koncích, takzvaných histonových ocasech, různými způsoby. Acetylace histonů obecně vede k uvolnění chromatinu a aktivaci genové exprese, zatímco deacetylace má opačný efekt. Metylace histonů může mít v závislosti na konkrétním místě a rozsahu modifikace jak aktivační, tak represivní účinky. Fosfory lace, ubikvitinace a sumoylace histonů jsou dalšími modifikacemi, které tvoří komplexní systém regulace, jenž někteří vědci nazývají histonovým kódem.
Třetím významným epigenetickým mechanismem je regulace pomocí nekódujících RNA molekul. Malé interferující RNA, mikroRNA a dlouhé nekódující RNA mohou ovlivňovat expresi genů na různých úrovních, od transkripce přes posttranskripční modifikace až po translaci. Tyto molekuly tvoří složité regulační sítě, jejichž plné pochopení je stále předmětem intenzivního výzkumu.
Epigenetické změny nejsou náhodné. Jsou ovlivňovány celou řadou faktorů vnějšího prostředí, jako jsou výživa, stres, expozice chemickým látkám, fyzická aktivita nebo sociální interakce. Slavný příklad z oblasti výzkumu epigenetiky představují studie na jednovaječných dvojčatech. Přestože tato dvojčata sdílejí identickou genetickou výbavu, jejich epigenomy se s přibývajícím věkem stále více liší, a to v závislosti na jejich odlišných životních zkušenostech a prostředí, ve kterém žijí. Tato pozorování přesvědčivě dokládají, že epigenetické modifikace jsou dynamické a mohou se měnit v průběhu života jedince.
Zvláštní pozornost si zaslouží fenomén transgeneračního epigenetického dědění. Tradičně se předpokládalo, že epigenetické modifikace jsou při tvorbě pohlavních buněk a po oplodnění zcela vymazány a znovu nastaveny. Výzkumy posledních let však ukázaly, že tento proces takzvaného epigenetického reprogramování není vždy úplný a některé epigenetické značky mohou přetrvat a být přeneseny na potomstvo. Nejznámějším příkladem je takzvaný holandský hladový winter z let 1944 až 1945, kdy bylo prokázáno, že podvýživa matek měla měřitelné epigenetické a zdravotní dopady nejen na jejich děti, ale i na vnoučata.
V kontextu genetiky jako vědecké disciplíny a v rámci výuky genetiky na středních a vysokých školách je epigenetika tématem, které výrazně obohacuje tradiční mendelovský pohled na dědičnost. Ukazuje, že vztah mezi genotypem a fenotypem je mnohem komplexnější, než se dříve předpokládalo, a že prostředí může ovlivňovat projev genetické informace způsoby, které mohou být za určitých okolností přenášeny na další generace. Pochopení epigenetických mechanismů má zásadní význam nejen pro základní výzkum, ale také pro medicínu, kde epigenetické změny hrají důležitou roli v rozvoji nádorových onemocnění, neurologických poruch, metabolických chorob a mnoha dalších patologických stavů. Epigenetická terapie, tedy cílená modifikace epigenetických značek pomocí farmakologických látek, představuje slibnou oblast moderní medicíny, která otevírá nové možnosti léčby dosud obtížně léčitelných onemocnění.
Genetické choroby představují rozsáhlou skupinu onemocnění, jejichž příčina spočívá v poškození nebo abnormalitě genetické informace uložené v DNA. Pochopení těchto chorob vyžaduje solidní základ v oblasti molekulární biologie a klasické genetiky, přičemž každá moderní genetika učebnice věnuje tomuto tématu značnou pozornost. Genetické choroby mohou být způsobeny mutacemi v jediném genu, chromozomálními aberacemi nebo komplexními interakcemi mezi mnoha geny a faktory prostředí.
Mezi nejlépe prostudované monogenní choroby patří například cystická fibróza, která vzniká mutací genu CFTR na sedmém chromozomu. Tato choroba postihuje funkci chloridových kanálů v buněčných membránách, což vede k hromadění hustého hlenu v dýchacích cestách a trávicím traktu. Diagnostika cystické fibrózy se opírá o novorozenecký screening, potní test a molekulárně genetické vyšetření, při němž se identifikují konkrétní mutace v genu CFTR. Moderní terapie zahrnuje nejen symptomatickou léčbu, ale v posledních letech také modulátory CFTR proteinu, jako jsou ivakaftor, lumakaftor nebo tezakaftor, které přímo ovlivňují funkci poškozeného proteinu a představují průlom v léčbě tohoto závažného onemocnění.

Chromozomální aberace tvoří další velkou skupinu genetických chorob. Downův syndrom, způsobený trizomií 21. chromozomu, je nejznámějším příkladem numerické chromozomální aberace. Diagnostika se provádí prenatálně pomocí biochemického screeningu v prvním trimestru těhotenství, ultrazvukového vyšetření a v případě potřeby invazivními metodami, jako je amniocentéza nebo odběr choriových klků, při nichž se provádí karyotypizace plodu. V posledních letech se rozšířilo neinvazivní prenatální testování, známé jako NIPT, které analyzuje volnou fetální DNA v mateřské krvi a umožňuje detekci chromozomálních aberací s vysokou senzitivitou a specificitou.
Diagnostické možnosti v oblasti genetických chorob se v průběhu posledních desetiletí dramaticky rozšířily. Sekvenování nové generace, označované jako NGS, umožňuje analyzovat celý exom nebo dokonce celý genom pacienta a odhalit i vzácné mutace, které by dříve zůstaly nedetekované. Tato technologie se stala nepostradatelným nástrojem při diagnostice vzácných genetických chorob, kde pacient prošel dlouhou diagnostickou odysejí bez stanovení diagnózy. Panelové sekvenování cílené na skupiny genů spojených s určitým klinickým fenotypem pak představuje kompromis mezi cenou, časem a diagnostickým výtěžkem.
Fenylketonurie je dalším klasickým příkladem genetické choroby, u níž včasná diagnostika a léčba zásadně ovlivňují prognózu pacienta. Tato autosomálně recesivní choroba způsobená mutacemi v genu PAH vede k hromadění fenylalaninu v organismu, což bez léčby způsobuje těžkou mentální retardaci. Díky novorozeneckému screeningu, který se v České republice provádí od sedmdesátých let minulého století, je možné zahájit dietní léčbu s omezením fenylalaninu ještě před rozvojem neurologického poškození. Moderní terapeutické přístupy zahrnují také podávání sapropterinu, kofaktoru enzymu fenylalanin hydroxylázy, u pacientů s určitými typy mutací.
Genová terapie představuje jeden z nejslibnějších směrů v léčbě genetických chorob. Principem je zavedení funkční kopie genu do buněk pacienta, čímž se kompenzuje defektní nebo chybějící funkce původního genu. Vektory pro přenos genetické informace mohou být virové, nejčastěji adeno-asociované viry nebo lentivirové vektory, nebo nevirové. Spinální muskulární atrofie, způsobená mutacemi v genu SMN1, se dnes léčí přípravkem onasemnogen abeparvovec, který jednorázovou intravenózní infuzí dodá funkční kopii genu SMN1 do buněk míchy. Výsledky klinických studií jsou pozoruhodné, zejména u pacientů léčených v raném věku před rozvojem závažného neurologického poškození.
CRISPR-Cas9 technologie otevřela zcela nové možnosti přesného editování genomu. Tato metoda využívá bakteriální obranný systém k cílenému přestřižení DNA na přesně definovaném místě, po němž může dojít k opravě mutace nebo vnesení žádoucí změny. V klinické praxi se tato technologie začíná uplatňovat například v léčbě srpkovité anémie a beta-talasémie, kde editace genu v kmenových buňkách krvetvorby vede k reaktivaci fetálního hemoglobinu, který kompenzuje defektní dospělý hemoglobin. Etické aspekty editace zárodečné linie, tedy změn, které by se dědily na potomstvo, jsou předmětem intenzivní odborné i společenské diskuse a v současnosti jsou ve většině zemí zakázány.
Farmakogenetika, studující vliv genetické variability na odpověď organismu na léky, se stala nedílnou součástí personalizované medicíny. Polymorfismy v genech kódujících enzymy metabolizující léčiva, jako jsou geny rodiny CYP450, určují, zda bude pacient rychlý nebo pomalý metabolizátor daného léku, což ovlivňuje jeho účinnost i bezpečnost. Znalost genetického profilu pacienta tak umožňuje individualizovat dávkování léků a předcházet nežádoucím účinkům nebo terapeutickým selháním.
Genetické poradenství hraje klíčovou roli v péči o rodiny postižené genetickými chorobami. Genetický poradce pomáhá rodinám pochopit podstatu onemocnění, způsob dědičnosti, riziko pro potomky a dostupné možnosti diagnostiky a léčby. Preimplantační genetická diagnostika umožňuje párům s rizikem přenosu genetické choroby vybrat zdravá embrya při procesu in vitro fertilizace ještě před jejich přenosem do dělohy, čímž se předchází narození postiženého dítěte. Tato metoda je dostupná pro stále širší spektrum genetických chorob a chromozomálních aberací.
Genetika jako věda přináší do moderní medicíny a biologie obrovské množství poznatků, které mají přímý dopad na životy jednotlivců i celých společností. S každým novým objevem v oblasti genomiky, sekvenování DNA nebo genetického testování se však vynořují otázky, které přesahují rámec čistě vědeckého bádání a dotýkají se hlubokých etických, filozofických a společenských hodnot. Tyto otázky nelze ignorovat, a proto jim moderní genetika učebnice věnují stále více prostoru, protože budoucí vědci, lékaři a genetičtí poradci musí být připraveni na složitost morálních dilemat, která jejich práce přináší.
Jedním z nejzásadnějších etických problémů je otázka genetického soukromí. Každý člověk nese ve svém genomu informace, které jsou jedinečné a zároveň sdílené s jeho biologickými příbuznými. Pokud si jedinec nechá provést genetické testování, výsledky mohou odhalit informace týkající se nejen jeho samotného, ale i jeho rodičů, sourozenců nebo dětí. Vzniká tak napětí mezi právem jednotlivce na informace o svém zdravotním stavu a právem příbuzných na ochranu jejich soukromí. Kdo má právo rozhodovat o tom, zda budou tyto informace sdíleny? A co se stane, když genetické informace skončí v rukou zaměstnavatelů nebo pojišťoven?
Genetická diskriminace představuje reálné riziko, které moderní legislativa v mnoha zemích teprve začíná řešit. Existují obavy, že lidé s prokázanou genetickou predispozicí k určitým onemocněním mohou být znevýhodněni při získávání zaměstnání nebo zdravotního pojištění. Ačkoli některé státy přijaly zákony zakazující genetickou diskriminaci, jejich vymahatelnost a rozsah ochrany se výrazně liší. Genetika učebnice proto zdůrazňují, že studenti musí chápat právní rámec genetického testování v kontextu konkrétní země, ale zároveň si uvědomovat jeho globální přesah.
Dalším klíčovým tématem je prenatální genetické testování a jeho etické důsledky. Možnost odhalit genetické abnormality plodu ještě před narozením přináší rodičům informace, které mohou zásadně ovlivnit jejich rozhodnutí o pokračování těhotenství. Tato situace vyvolává hluboké etické debaty o hodnotě lidského života, o právu na život jedince s postižením a o hranicích rodičovské autonomie. Někteří etici upozorňují, že rozšíření prenatálního testování může vést k selektivnímu přístupu k životu, který je v přímém rozporu s hodnotami inkluzivní společnosti. Jiní naopak argumentují, že rodiče mají právo na co nejúplnější informace pro svá rozhodnutí.
Preimplantační genetická diagnostika, která umožňuje výběr embryí před jejich implantací při umělém oplodnění, posouvá tyto otázky ještě dál. Hranice mezi léčbou genetických onemocnění a výběrem žádoucích vlastností potomka se stává stále tenčí. Pojem „designer babies, tedy geneticky upravené děti s předem vybranými vlastnostmi, přestává být pouhou sci-fi fantazií a stává se reálnou možností, kterou věda musí zodpovědně reflektovat. Kde leží hranice mezi terapeutickým využitím genetiky a eugenickými tendencemi? Tato otázka je jednou z nejpalčivějších, s nimiž se současná genetika potýká.
Eugenika jako historický fenomén zanechala v dějinách hluboké jizvy. Programy nucené sterilizace, které byly v první polovině dvacátého století realizovány v různých zemích světa, včetně demokratických, jsou dnes považovány za hrubé porušení lidských práv. Moderní genetika se musí důsledně distancovat od jakýchkoli eugenických tendencí, přičemž zároveň musí být schopna nabízet genetické poradenství způsobem, který respektuje autonomii jedince a nepodléhá paternalistickým přístupům. Genetika učebnice proto věnují zvláštní pozornost historickému kontextu eugeniky jako varování před zneužitím vědeckých poznatků.
Genetické poradenství samo o sobě představuje oblast plnou etických výzev. Genetický poradce musí být schopen sdělovat složité a často velmi závažné informace způsobem, který je srozumitelný, ale zároveň respektuje emocionální stav klienta. Princip nedirektivního poradenství, který je dnes považován za standard, předpokládá, že poradce neprosazuje vlastní hodnotové soudy a ponechává rozhodnutí plně na klientovi. V praxi je však tento princip obtížně udržitelný, protože způsob, jakým jsou informace prezentovány, vždy do určité míry ovlivňuje rozhodování.
Výzkum lidského genomu přináší další etické dilema v podobě nakládání s genetickými daty. Velké databáze genetických informací, které jsou nezbytné pro výzkum komplexních onemocnění, musí být budovány s maximálním ohledem na soukromí dárců. Anonymizace dat, informovaný souhlas a transparentnost výzkumných záměrů jsou základními požadavky, jejichž naplnění však není vždy snadné. Zvláště problematická je situace, kdy jsou genetická data sdílena mezi výzkumnými institucemi v různých zemích s odlišnými právními standardy ochrany osobních údajů.
Otázka vlastnictví genetických informací a patentování genů je dalším kontroverzním tématem. Může být gen patentován jako vynález, nebo jde o součást přirozeného dědictví lidstva, které by mělo být volně dostupné? Spory o patenty na geny spojené s rizikem rakoviny prsu ukázaly, jak obrovské ekonomické a etické důsledky mohou mít tato rozhodnutí pro přístup pacientů k diagnostice a léčbě. Genetika učebnice nemohou tyto otázky obcházet, protože formují nejen vědecké myšlení, ale i schopnost studentů kriticky hodnotit společenský dopad vědy.
Genetické testování přímým přístupem spotřebitele, tedy testování bez zprostředkování lékařem, se v posledních letech masivně rozšířilo. Komerční společnosti nabízejí analýzu DNA za relativně nízké ceny a slibují informace o původu, zdravotních predispozicích nebo vloženích pro různé vlastnosti. Tato demokratizace přístupu ke genetickým informacím má nepochybně své výhody, ale přináší i závažná rizika. Lidé bez odborného vzdělání mohou špatně interpretovat výsledky, podlehnout zbytečné úzkosti nebo naopak falešnému pocitu bezpečí. Chybí jim kontext, který by jim pomohl pochopit, že genetická predispozice neznamená osud.

Celá oblast etiky genetického výzkumu a testování vyžaduje interdisciplinární přístup, který propojuje poznatky genetiky, medicíny, filozofie, práva a sociologie. Budoucí generace vědců a lékařů musí být vybavena nejen odbornými znalostmi, ale i schopností etické reflexe, která jim umožní zodpovědně nakládat s mocí, jež jim genetika dává do rukou. Genetika učebnice proto stále více integrují etické kapitoly jako nedílnou součást vědeckého vzdělání, protože věda bez svědomí může přinést více škody než užitku.
Publikováno: 18. 01. 2026
Kategorie: Genetika


